
Echinodermata
Échinodermes

| Empire | Eukaryota |
|---|---|
| Règne | Animalia |
| Sous-règne | Eumetazoa |
| Clade | Bilateria |
| Clade | Nephrozoa |
| Super-embr. | Deuterostomia |
| Clade | Ambulacraria |
Sous-embranchements de rang inférieur
Les échinodermes (Echinodermata) sont un embranchement d'animaux marins benthiques présents à toutes les profondeurs océaniques. Les environ 6 500 espèces[2] sont toutes exclusivement marines : on ne connaît aucune espèce terrestre ni d'eau douce. Elles sont actuellement regroupées en cinq classes : les étoiles de mer, les ophiures, les oursins, les concombres de mer et les lys de mer.
Les représentants de ce groupe possèdent des caractéristiques uniques dans le monde animal. Les principales sont une symétrie générale pentaradiée (bien qu'ils restent biologiquement des bilatériens[3]), un squelette externe constitué de plaques osseuses articulées et percées de nombreuses canalicules, et la présence d'un système aquifère. Dans la biodiversité de l'évolution, ils constituent un groupe deutérostomien proche des chordés.
Outre les espèces actuelles, plusieurs milliers d'espèces éteintes d'échinodermes sont recensées par les paléontologues, leur squelette très calcifié permettant généralement une bonne fossilisation. Leurs plus anciennes traces fossiles remontent au Cambrien[4].
Caractéristiques morphologiques[modifier | modifier le code]

Les échinodermes sont des animaux (métazoaires triploblastiques) caractérisés par leur endosquelette dermique (structure appelée « stéréome ») constitué de monocristaux de calcite monoaxiaux[5], formant des spicules (holothuries) ou des plaques, qui peuvent être mobiles (étoiles de mer, ophiures, crinoïdes) ou soudées (oursins, dans ce cas on appelle le squelette le « test »)[6]. De nombreuses espèces sont également hérissées de piquants[4]. Ces pièces squelettiques, quand elles ne sont pas soudées, sont tenues entre elles par un réseau de ligaments composés majoritairement de collagène (plus précisément « MCT », mutable collagenous tissue), dont la rigidité est commandée par influx nerveux ; cela permet aux échinodermes de rigidifier ou de ramollir leur corps à volonté, notamment pour maintenir des positions contractées sans effort musculaire ; cela est particulièrement le cas pour les holothuries, au corps peu calcifié[5].
La caractéristique la plus frappante est la symétrie pentaradiée (symétrie centrale d'ordre 5) pour la plupart des structures de la forme adulte[3] : cela est particulièrement visible sur les étoiles de mer par exemple, mais ce caractère n'est pas toujours aussi manifeste chez toutes les espèces, notamment les concombres de mer ou les oursins irréguliers[3]. Toutefois, leur symétrie interne et les formes embryonnaires, à symétrie clairement bilatérale[7], les rangent aux côtés des chordés dans le groupe des deutérostomiens : ils partagent d'ailleurs avec eux bon nombre de caractéristiques biochimiques[4]. Cette symétrie pentaradiaire est donc secondaire[3], pas toujours complète et ne concerne pas certains organes comme le madréporite, souvent unique.
En plus de se retrouver dans la morphologie générale, leur symétrie pentaradiée se retrouve également dans l'ensemble de l'organisation du corps, depuis le système nerveux jusqu'au squelette calcifié constitué de plaques et d'épines qui entourent et protègent l'intérieur du corps[7]. L'animal ne présente donc généralement pas de face « ventrale » et « dorsale » (typiques des bilatériens chordés) mais une face orale (où se situe la bouche) et une face aborale (où se trouvent généralement l'anus, l'appareil respiratoire et les gonades, ou le pied chez les crinoïdes fixés). Ils n'ont donc pas non plus de « tête » : les systèmes digestif, circulatoire et nerveux sont tous répartis en cinq branches symétriques ramifiées parcourant le corps[8]. Ces 5 éléments radiaires sont appelés « rayons » (comme les bras des étoiles de mer), et peuvent éventuellement être ramifiés (notamment chez les crinoïdes ou les ophiures gorgonocéphales). Ces rayons se développent depuis des plaques oculaires, d'où apparaissent des plaques ambulacraires sur deux rangées alternativement. Ce phénomène est bien visible sur le test des oursins[9].
Leur système vasculaire est réduit et relativement ouvert, sans organe pulsatile[7]. Ils disposent en plus d'un système aquifère propre à ce groupe, qui assure quelques-unes des fonctions du sang et quelques autres : échanges gazeux, locomotion (par jeu de pression différentielle dans les podia) et nutrition ; il est alimenté par un organe filtreur appelé madréporite[10]. Le système digestif est complet et parfaitement fonctionnel, proche de celui des chordés[11]. Le système nerveux est constitué d'un anneau nerveux autour de la bouche (anneau périoral) d'où partent des nerfs selon une structure radiaire en 5 cordons nerveux (pas de système nerveux central)[4].
La larve est généralement pélagique, et l'individu mature benthique (sauf quelques rares crinoïdes et holothuries nageurs, dont une holothurie pélagique). Ils peuvent être sessiles (comme les crinoïdes pédonculés) ou vagiles : certaines ophiures et étoiles de mer carnivores sont même capables de mouvements assez rapides[12].
Sur le plan développemental, une étude de génétique du développement a montré, sur la base d'une cartographie de l'expression des gènes développementaux, que « du point de vue de la structuration de l'ectoderme, les échinodermes sont des animaux principalement constitués d'une tête », n'exprimant nulle part les gènes typiques des régions postérieures[13].
-
 Une étoile de mer.
Une étoile de mer. -
 Une ophiure.
Une ophiure. -
 Un crinoïde.
Un crinoïde. -
 Un oursin.
Un oursin. -
 Un concombre de mer.
Un concombre de mer.





Écologie et comportement[modifier | modifier le code]

Habitat[modifier | modifier le code]
Les échinodermes sont exclusivement marins : on n'en connaît aucune trace en eau douce, y compris parmi les formes fossiles. Ils peuplent pratiquement toutes les mers ouvertes du globe (ils ont cependant disparu des mers fermées comme la Caspienne), depuis la surface jusqu'aux abysses, et de l'équateur jusqu'aux mers subglaciales[4]. Ils font partie des groupes d'animaux supérieurs les plus représentés dans les profondeurs abyssales, plusieurs espèces de chaque classe étant présentes au-delà de 8 000 m de profondeur, et certaines espèces d'holothuries jusqu'à plus de 10 000 mètres[14]. Ils constituent une part très importante de la faune benthique, et jouent donc un rôle majeur dans les processus biologiques de ces milieux, notamment en ce qui concerne le recyclage des déchets par leurs régimes détritivores ou filtreurs.
On compte par exemple une centaine d'espèces en Méditerranée occidentale[15], réparties en 22 étoiles de mer, 23 ophiures, 2 crinoïdes, 22 oursins et 31 holothuries, toutes profondeurs confondues.
Reproduction[modifier | modifier le code]
Les sexes sont séparés et la reproduction gonochorique, mais il n'existe pas de dimorphisme sexuel. La disposition de l'appareil reproducteur varie selon les espèces : à l'apex (sommet) des oursins et de certaines étoiles, au niveau des pinnules chez les crinoïdes, dans la cavité centrale des holothuries, au niveau des bras pour les ophiures et la plupart des étoiles[8]. La plupart des espèces émettent une grande quantité de gamètes (jusqu'à plusieurs millions d’œufs peuvent être fécondés en une seule reproduction) : le sperme et les ovocytes sont libérés dans le milieu de manière synchrone grâce à un signal phéromonal, et la fécondation a ensuite lieu au hasard dans l'eau. Les larves évoluent ensuite parmi le plancton pendant plusieurs semaines (où elles sont la proie de nombreux planctonivores), subissant des métamorphoses caractéristiques des différentes classes (mais toutes sont encore à symétrie bilatérale)[16]. Arrivées au terme de leur dernier cycle larvaire, elles se rapprochent du fond jusqu'à s'y poser pour entamer leur dernière métamorphose en juvénile, et commencer leur cycle de vie[8].
Les échinodermes, et particulièrement les oursins, ont été très utiles dans la compréhension du phénomène de fécondation, notamment en ce qui concerne les mouvements ioniques lors de la pénétration du spermatozoïde[17],[18].
Certaines espèces ont cependant développé des modes de reproduction particuliers : absence de stade planctonique[19], autofécondation, reproduction par division (chez les étoiles et les holothuries)[20], etc.
-
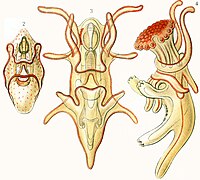
-
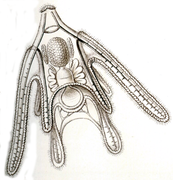 Larve ophioplutée d’ophiure.
Larve ophioplutée d’ophiure. -
 Larve brachiolaire d'étoile de mer.
Larve brachiolaire d'étoile de mer. -
 Larve échinoplutée d'oursin.
Larve échinoplutée d'oursin. -
 Larve auriculaire d'holothurie.
Larve auriculaire d'holothurie. -
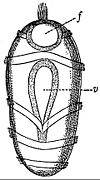 Larve doliolaire d'un crinoïde.
Larve doliolaire d'un crinoïde.






Locomotion[modifier | modifier le code]

Presque tous les échinodermes sont benthiques : cela signifie qu'ils vivent posés sur le fond, à l'exception de certaines rares espèces d'holothuries capables de vivre en pleine eau (comme Pelagothuria natatrix, mais plusieurs autres espèces peuvent nager un moment[21], tout comme certaines comatules[22]).
De nombreuses espèces sont « sessiles », ce qui signifie qu'elles vivent fixées au substrat, où elles se nourrissent en filtrant l'eau : c'est le cas de nombreux crinoïdes et de certaines holothuries (et dans une moindre mesure de certaines espèces des trois autres classes). À l'inverse, les échinodermes capables de se déplacer sont dits « vagiles », et certaines étoiles de mer et ophiures sont capables de mouvements relativement rapides[12].
Pour se déplacer et adhérer à des surfaces dures, les oursins, les étoiles de mer et les holothuries sont pourvus de minuscules « pieds ambulacraires » en forme de tubes mous, munis à leur extrémité de cellules collantes et appelés podia[4],[23]. Ceux-ci peuvent développer une force d'adhérence extrêmement efficace[24], comme chez l'oursin-tortue Colobocentrotus atratus[25].
Alimentation[modifier | modifier le code]

Les différentes espèces d'échinodermes ont adopté une grande variété de régimes alimentaires. Ils peuvent être des prédateurs carnivores (principalement les étoiles de mer, mais aussi certains oursins et ophiures), charognards (étoiles, oursins et ophiures), brouteurs herbivores (la plupart des oursins), dépositivores (oursins, holothuries, ophiures) fouisseurs (oursins irréguliers) ou encore filtreurs (crinoïdes, holothuries). Leur bouche est généralement très développée et adaptée au régime, avec par exemple des tentacules digités ou ramifiés en fractales pour filtrer le plancton ou le sédiment chez les holothuries[8], un estomac pouvant être dévaginé pour assurer la digestion externe de grosses proies chez de nombreuses étoiles de mer[8], ou encore une puissante mâchoire dentée (appelée « lanterne d'Aristote ») chez les oursins[26].
On constate que le régime suspensivore sessile, qui semble être le plus ancien dans ce groupe, est une caractéristique ponctuellement récurrente chez toutes les classes malgré leur diversité : des ordres d'étoiles de mer (comme les Brisingida), d'ophiures (comme les Euryalida), d'holothuries (Dendrochirotida) et même certains oursins (Dendraster excentricus[27], peut-être aussi Dermechinus horridus[28]) sont revenus à ce mode de vie primitif qui est celui des crinoïdes (et avant eux de groupes archaïques comme les Eocrinoidea), développant des convergences morphologiques (principalement de longs bras rétractiles ramifiés en fractales ciliées).
Dans les groupes carnivores actifs, la prédation intra-embranchement n'est pas rare : ainsi de nombreuses étoiles de mer se nourrissent principalement d'autres échinodermes, comme les Solaster[29]. Cette grande diversité en fait des animaux extrêmement importants dans les cycles biologiques marins, présents à tous les niveaux trophiques en grande abondance. Ainsi, leurs fluctuations de population sont susceptibles d'avoir des effets importants sur tout l'écosystème.
Respiration[modifier | modifier le code]
La respiration est passive chez la plupart des échinodermes, et s'effectue le plus souvent à travers la peau, et notamment au niveau du système aquifère. Elle est aussi assurée en partie de manière directe par les podia chez les espèces qui en sont pourvues, mais les espèces massives ayant de gros besoins sont souvent pourvues d'organes respiratoires plus spécialisés : organes arborescents chez certains ordres d'holothuries, branchies chez certains oursins réguliers (Echinacea), papules respiratoires chez certaines étoiles de mer, bourses branchiales chez les ophiures[8].
Records[modifier | modifier le code]

Le plus long échinoderme connu est sans conteste l'holothurie serpentiforme Synapta maculata, qui peut atteindre 3 m de long, pour seulement quelques centimètres de large (et un poids très limité, sa densité étant à peine supérieure à celle de l'eau)[30]. Les échinodermes les plus lourds atteignent 6 kg : ce poids a été mesuré chez certaines étoiles du genre Thromidia[31] et l'holothurie géante Thelenota anax[32].
L'échinoderme vivant le plus abyssal est probablement l'holothurie Myriotrochus bruuni collectée à 10 687 mètres de profondeur, dans la fosse des Mariannes[14].
Peu d'échinodermes sont dangereux pour les humains : on ne connaît qu'une ou deux étoiles de mer venimeuses (notamment Acanthaster planci), et quelques familles d'oursins (Diadematidae, Echinothuriidae et Toxopneustidae) : seule une espèce d'oursin semble pouvoir être occasionnellement mortelle pour l'Homme, l'oursin-fleur Toxopneustes pileolus[33].
Il est difficile d'établir l'espèce d'échinoderme la plus répandue à l'échelle mondiale, d'autant plus qu'il conviendrait de distinguer entre effectif (valorisant les espèces minuscules) et biomasse ; cependant l'ophiure Ophiactis savignyi, très généraliste et capable de se reproduire par voie asexuée, est signalée dans quasiment toutes les mers du globe[34]. Plus visible, l'échinoderme littoral le plus facilement rencontré est probablement l'oursin tropical Echinometra mathaei, extrêmement abondant de la Mer Rouge à Hawaï[35]. Cependant, plusieurs espèces abyssales ont sans doute une répartition et des effectifs encore plus importants, les surfaces exploitables étant très supérieures.
Enfin, on ne connaît à l'heure actuelle qu'une seule espèce pélagique, c'est-à-dire vivant en permanence dans la colonne d'eau sans jamais se poser sur le fond : l'énigmatique holothurie Pelagothuria natatrix[21]. En dehors de cette espèce très particulière, seule une poignée d'holothuries et de crinoïdes sont capables de nager.
Origine phylogénétique et registre paléontologique[modifier | modifier le code]
Les échinodermes sont un groupe extrêmement ancien, et dont l'histoire évolutive est relativement bien documentée grâce à leur squelette calcaire très minéralisé, qui permet souvent une excellente fossilisation[36].
Les plus anciens fossiles attestés d'échinodermes remontent à -525 millions d'années (Cambrien inférieur)[4], mais d'autres fossiles leur ressemblant pourraient être plus vieux encore[5].
Les échinodermes dériveraient de formes à symétrie bilatérale, les hétérostèles. La symétrie pentaradiée semble ne pas s'être imposée immédiatement, une forme précambrienne d'Ediacara (Tribrachidium) est en effet triradiée, mais son appartenance aux échinodermes est très controversée ; les homalozoa du Cambrien semblent quant à eux bilatériens, mais leur inclusion parmi les échinodermes est elle aussi parfois remise en question, le groupe étant accusé par certains d'être paraphylétique[37]. Les plus anciens animaux à symétrie pentaradiée sont les Arkarua[5], trouvés à l'état de traces dans des sédiments de l'édiacarien (vers -560 millions d'années), mais leur squelette ne semble pas structuré par un stéréome, trait normalement caractéristique des échinodermes (et ne connaissant pas d'autre exception connue, contrairement à la symétrie pentaradiaire). Le taxon des carpoïdes (rangé dans le groupe des mitrales au sein des homalozoa) est lui aussi encore difficile à situer : il s'agit d'une forme étrange de l'Ordovicien que quelques scientifiques classent parmi les chordés primitifs[37]. Les formes certaines les plus anciennes sont des organismes sessiles suspensivores et comprennent les éocrinoïdes, les cystoïdes et les blastoïdes, qui furent très abondants au Paléozoïque mais disparurent respectivement au Silurien, au Carbonifère et lors de l'extinction Permien-Trias.
Tous les spécimens du Mésozoïque et du Cénozoïque font partie des cinq classes actuelles, mais le registre Paléozoïque contient des animaux appartenant à d'autres classes, éteintes (jusqu'à 15)[5],[36]. Plusieurs clades majeurs d'échinodermes sont ainsi complètement éteints : les homalozoa (animaux du Cambrien à symétrie bilatérale pourvus d'un bras unique appelé aulacophore), les Helicoplacoidea (échinodermes du Cambrien en forme de pelotes), les Cyclocystoidea (en forme de tambourins), les Edrioasteroidea (petits animaux sessiles étoilés), les Somasteroidea (premiers animaux mobiles en forme d'étoile), les Ophiocistioidea (sortes d'oursins sans piquants munis de longs bras), les Eocrinoidea (ressemblant aux cinoïdes mais sans tige différenciée), les blastoidea (en forme de noix pédonculée) ou encore les Cystoidea (en forme de massues pédonculées)[4]. Aucun de ces groupes n'a passé la crise Permien-Trias, qui mit fin à l'ère Paléozoïque[36].
Par la suite, les échinodermes survivants à cette crise ont cependant continué de laisser d'abondantes traces, en particulier les groupes des crinoïdes et des oursins, qui sont présents dès l'Ordovicien et abondants à partir du Carbonifère, et dont les corps durs permettent une excellente fossilisation (ce qui n'est pas le cas des holothuries, par exemple[38]). Au total 13 000 espèces éteintes ont été identifiées (contre environ 7 000 vivantes aujourd'hui)[5].
| Nom | Apparition | Disparition | Remarques |
|---|---|---|---|
| Arkarua | -635 millions d'années (Édiacarien) | -542 millions d'années (Cambrien) | Statut encore incertain, pas de stéréome |
| Helicoplacoidea | -530 Ma (Cambrien) | -516 Ma (Cambrien) | Symétrie pas encore affirmée |
| Eocrinoidea | -530 Ma (Cambrien) | -298 Ma (Silurien) | |
| Homalozoa | -510 Ma (Cambrien) | -300 Ma (Carbonifère) | statut et cohérence discutés, symétrie bilatérale |
| Cystoidea | -513 Ma (Cambrien) | -392 Ma (Carbonifère) | |
| Edrioasteroidea | -516 Ma (Cambrien) | -272 Ma (Permien) | |
| Somasteroidea | -490 Ma (Ordovicien) | -350 Ma (Dévonien) | |
| Cyclocystoidea | -471 Ma (Ordovicien) | -379 Ma (Dévonien) | |
| Blastoidea | -488 Ma (Ordovicien) | -252 Ma (Permien) | |
| Rhombifera | -460 (Ordovicien) | -390 Ma (Dévonien) | |
| Ophiocistioidea | -475 (Ordovicien) | -254 Ma (Permien) | |
| Crinoidea | -470 Ma (Ordovicien) | actuel | Crinoïdes et comatules |
| Echinoidea | -450 Ma (Ordovicien) | actuel | Oursins |
| Asteroidea | -440 Ma (Ordovicien) | actuel | Étoiles de mer |
| Ophiuroidea | -440 Ma (Ordovicien) | actuel | Ophiures |
| Holothuroidea | inconnu (fossilisation rare) | actuel | Holothuries |
-
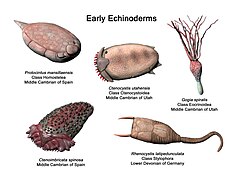 Représentation de quelques échinodermes paléozoïques par le paléoartiste Nobu Tamura.
Représentation de quelques échinodermes paléozoïques par le paléoartiste Nobu Tamura. -
 Tribrachidium heraldicum, possible forme ancestrale des échinodermes.
Tribrachidium heraldicum, possible forme ancestrale des échinodermes. -
 Fossile de Streptaster vorticellatus (Edrioasteroidea).
Fossile de Streptaster vorticellatus (Edrioasteroidea). -
 Dessin d'Ophiocistioidea.
Dessin d'Ophiocistioidea. -
 Fossile d'éocrinoïde (Gogia spiralis).
Fossile d'éocrinoïde (Gogia spiralis). -
 Un éocrinoïde dessiné par Ernst Haeckel (1904).
Un éocrinoïde dessiné par Ernst Haeckel (1904). -
 Un homalozoaire par Haeckel (1904).
Un homalozoaire par Haeckel (1904). -
 Des cystoïdes (ibid).
Des cystoïdes (ibid). -
 Des blastoïdes (ibid).
Des blastoïdes (ibid). -
 Fossiles de crinoïdes.
Fossiles de crinoïdes. -
 Fossile du somastéroïde (Stuertzia leptosoma).
Fossile du somastéroïde (Stuertzia leptosoma). -
 Fossile de l'oursin Archaeocidaris brownwoodensis.
Fossile de l'oursin Archaeocidaris brownwoodensis. -
 Fossile d'Homalozoaire (Cothurnocystis elizae).
Fossile d'Homalozoaire (Cothurnocystis elizae). -
 Fossile du rhombifère (Pleurocystites squamosus).
Fossile du rhombifère (Pleurocystites squamosus).


_from_the_Bellevue_Formation_(Upper_Ordovician)_at_the_Maysville_West_roadcut_of_northern_Kentucky,_USA.jpg)











Classification[modifier | modifier le code]
Histoire scientifique[modifier | modifier le code]

L'un des plus anciens textes scientifiques concernant les échinodermes remonte à Aristote, dans son Histoire des animaux (vers -343) : il y décrit succinctement les étoiles de mer et plus longuement les oursins, et nomme un animal « holothurie » sans le décrire (il le rapproche des éponges car c'est un animal qui semble dépourvu de sens) ; ce nom sera conservé par la suite pour nommer les concombres de mer (classe holothuroidea). Le nom du philosophe sera quant à lui attribué à l'appareil masticateur des oursins, à travers une métaphore de son cru : la « lanterne d'Aristote »[39]. En Europe, Aristote demeure l'inspiration principale de la plupart des traités d'histoire naturelle de l'antiquité et du Moyen Âge, et Pline l'Ancien s'en inspire ouvertement quand il aborde les mollusques (au rang desquels il compte les échinodermes) au livre IX de son Histoire naturelle (vers 77)[40]. Ces deux auteurs demeureront les références majeures pendant plus d'un millénaire et demi, repris jusque chez
Les scientifiques européens recommencent à s'intéresser aux échinodermes à partir de la Renaissance, et en 1553 Pierre Belon est le premier à proposer des affinités entre holothuries, étoiles de mer et oursins[41]. En 1554, Guillaume Rondelet corrige et enrichit les descriptions antiques, et commence lui aussi à noter chez certains « zoophytes » ce plan d'organisation « en rose »[42].
Les véritables progrès arrivent avec le siècle des Lumières : en 1751 les oursins, les holothuries et les étoiles de mer ont leur article dans l'Encyclopédie, mais sont encore dispersés et classés parmi les mollusques, avec quelques hésitations[43]. En 1758, Carl von Linné énumère de nombreuses espèces dans son Systema naturae, les plaçant chacune dans un seul genre (Echinus, Holothuria, etc.), toujours parmi les mollusques. C'est Jacob Theodor Klein qui le premier avait eu l'idée, en 1734, de regrouper les oursins non plus parmi les mollusques mais avec les étoiles de mer, concombres de mer, ophiures et crinoïdes, sous l'appellation d'« échinodermes » (sur le critère du plan d'organisation pentaradié) ; mais il faudra attendre que ses travaux soient poursuivis par Nathanael Gottfried Leske (1778) puis systématisés par Jean-Guillaume Bruguière en 1791 pour que le clade des échinodermes soit définitivement incorporé aux classifications scientifiques. En 1801, ce groupe est encore rapproché par Lamarck des cnidaires qui partagent un plan d'organisation radiaire (il parle de « radiaires échinodermes »), classification qui perdurera jusqu'à Louis Agassiz (dans les années 1830), avant que les données embryologiques démontrent l'éloignement de ces groupes. Au début du XIXe siècle, de nombreux explorateurs scientifiques décrivent en quelques décennies des dizaines de nouvelles espèces, qui furent rapidement divisées en nouveaux genres, puis familles et ordres. Dans les années 1880 sont publiés les épais volumes du Report Of The Scientific Results of the Exploring Voyage of H.M.S. Challenger during the years 1873-76 qui suivit l'expédition du Challenger, et permit la description d'une énorme quantité de nouveaux taxons, notamment en eaux profondes. Les premiers travaux de phylogénie scientifique apparaissent à partir du milieu du siècle (avec Louis Agassiz et Edouard Desor pour les oursins). Sous l'impulsion de Mortensen entre 1928 et 1951 (Monograph of Echinoidea), les critères squelettiques commencent à être préférés pour la classification des échinodermes (se fondant sur les plaques chez les étoiles de mer et ophiures, et les spicules chez les holothuries), permettant de réunir les classifications fossile et naturaliste[44]. En plus de Mortensen, plusieurs importantes monographies auront ainsi marqué la première moitié du XXe siècle, comme la Monograph of the Existing Crinoidea d'Austin H. Clark (à partir de 1915), ou encore les Starfish Monographs de Walter K. Fisher (1906-1940)[45].
La science des échinodermes est devenue un champ d'études consacré en 1963, avec l'organisation à Washington du premier Symposium on Physiology of Echinodermata (à l'origine une émanation du 16e International Congress of Zoology). À partir de 1968, la communauté des biologistes travaillant sur les échinodermes est fédérée par The Echinoderm Newsletter[46], à défaut d'une revue dédiée (contrairement à la plupart des autres embranchements du vivant), suivie par Echinoderm Studies de 1983 à 2001, puis le SPC Beche-de-mer Information Bulletin à partir de 1990 (plus spécialement consacré aux holothuries). Des publications ponctuelles semi-régulières permettent cependant de constituer des pierres de touche dans le progrès de la recherche, comme Echinoderms, Echinoderm Nutrition, ou Edible Sea Urchins[47]. En 1999, The Echinoid Directory est le premier site internet contenant une base de données de référence tenue à jour par des spécialistes, permettant notamment une homogénéisation de la taxinomie ; cette base de données est intégrée en 2008 au World Register of Marine Species, regroupant tous les échinodermes (et tous les autres règnes du vivant marin), qui constitue toujours la base de données en ligne de référence[47].
On connaît à l'heure actuelle environ 7 000 espèces vivantes d'échinodermes. Le groupe le plus diversifié est celui des ophiures avec près de 2 100 espèces, devant les étoiles de mer (autour de 1 900 espèces), puis les holothuries (1 250 espèces environ), les oursins (950 espèces) et les crinoïdes (600 espèces)[48]. Cependant, ce sont les oursins qui ont la plus grande diversité phylogénétique avec 16 ordres actuels (et plus de 70 familles, qui sont autant de plans d'organisation originaux), devant les étoiles (8 ordres), les holothuries (7), les ophiures (6 ordres) et les crinoïdes (4 ordres)[49].
Environ un millier d'espèces sont observables à l'œil nu dans le monde (1 137 espèces recensées sur le site collaboratif iNaturalist[50]), les autres étant au choix minuscules, cryptiques ou abyssales.
Classifications actuelles[modifier | modifier le code]

|
Selon World Register of Marine Species (28 août 2013)[51] :
|
Selon ITIS (28 août 2013)[52] :
|
Selon NCBI (28 août 2013)[53] :
|
Classification complète incluant les groupes fossiles, selon BioLib (5 janvier 2018)[54] :
- sous-embranchement Asterozoa Zittel, 1895
- classe Asteroidea (de Blainville, 1830)
- classe Ophiuroidea (Gray, 1840)
- classe Somasteroidea Spencer, 1951 †
- classe Stenuroidea Spencer, 1951 †
- famille Catervaparmasteridae Blake, 2000 †
- sous-embranchement Blastozoa Sprinkle, 1973 †
- classe Blastoidea Say, 1825 †
- classe Cystoidea von Buch, 1846 †
- classe Eocrinoidea Jaekel, 1899 †
- classe Parablastoidea †
- classe Paracrinoidea Regnéll, 1945 †
- classe Rhombifera Zittel, 1879 †
- sous-embranchement Crinozoa Matsumoto, 1929
- classe Crinoidea Miller, 1821
- classe Edrioasteroidea Billings, 1858 †
- sous-embranchement Echinozoa von Zittel 1895
- classe Cyclocystoidea Miller & Gurley, 1895 †
- classe Echinoidea Leske, 1778
- classe Edrioblastoidea Fay, 1962 †
- classe Helicoplacoidea †
- classe Holothuroidea (de Blainville, 1834)
- classe Ophiocistoidea Sollas, 1899 †
- sous-embranchement Homalozoa Whitehouse, 1941 †
- classe Ctenocystoidea Robison & Sprinkle, 1969 †
- classe Homoiostelea †
- classe Homostelea †
- classe Stylophora Gill & Caster, 1960 †
- genre Ctenoimbricata Zamora, Rahman & Smith, 2012 †
- genre Oryctoconus Colchen & Ubaghs, 1970 †
Place des échinodermes dans le règne animal[modifier | modifier le code]
- Les types d'organisation présentés ici sont des grades évolutifs ne correspondant généralement pas à des groupes monophylétiques, mais paraphylétiques (ne comportant pas tous les descendants d'un même ancêtre — exemple : les descendants d'ancêtres vermiformes ne sont pas tous aujourd'hui des vers, etc.).
- En jaune : les principales explosions radiatives.
Phylogénie des échinodermes d'après Telford et al. 2014[55] :
| Bilateria |
| ||||||||||||||||||||||||||||||||||||||||||||||||


Dans la culture[modifier | modifier le code]
Animaux exclusivement marins et sans tête différenciée, les échinodermes constituent un groupe biologique faiblement représenté dans la culture occidentale, car peu anthropomorphiques. Ils figurent cependant (avec une rigueur scientifique et orthographique limitée) dans le célèbre roman sous-marin de Jules Verne, Vingt mille lieues sous les mers : « Dans les échinodermes, remarquables par leur enveloppe épineuse, les astéries, les étoiles de mer, les pantacrines, les comatules, les astérophons, les oursins, les holoturies, etc., représentaient la collection complète des individus de ce groupe. »
Le chanteur français Gérard Manset a également composé une Ballade des échinodermes, sortie en 1994[56].
Les échinodermes sont aussi une source d'inspiration pour le biomimétisme et la science des biomatériaux[57].
Annexes[modifier | modifier le code]
Articles connexes[modifier | modifier le code]
Bibliographie[modifier | modifier le code]
- René Koehler, Faune de France : 1 : Échinodermes, Paris, Librairie de la Faculté des Sciences, (lire en ligne).
- (en) A.M. Clark et F.W.E. Rowe, Monograph of Shallow-water Indo-West Pacific Echinoderms, Londres, Trustees of the British Museum (Natural History), , 238 p. (lire en ligne).
- Alain Guille, Pierre Laboute et Jean-Louis Menou, Guide des étoiles de mer, oursins et autres échinodermes du lagon de Nouvelle-Calédonie, ORSTOM, , 244 p. (lire en ligne)
- Chantal Conand, Sonia Ribes-Beaudemoulin, Florence Trentin, Thierry Mulochau et Émilie Boissin, Oursins, étoiles de mer & autres échinodermes : Biodiversité de La Réunion, La Réunion, Les éditions du Cyclone, , 168 p. (ISBN 979-10-94397-04-6).
- (en) Maria Byrne et Timothy O'Hara, Australian echinoderms : Biology, Ecology and Evolution, CSIRO Publishing, (ISBN 978-1-4863-0763-0, lire en ligne).
- Frédéric Ducarme, Étoiles de mer, oursins et autres échinodermes de Mayotte et sa région, Les Naturalistes de Mayotte, , 336p (ISBN 978-2-9521543-5-2).
Références taxinomiques[modifier | modifier le code]
- (en) Référence WoRMS : Echinodermata Bruguière, 1791 [ex Klein, 1734] (+ liste espèces)
- (en) Référence Paleobiology Database : Echinodermata Klein 1754
- (fr + en) Référence ITIS : Echinodermata Klein, 1734
- (en) Référence Tree of Life Web Project : Echinodermata
- (en) Référence BioLib : Echinodermata Klein, 1734
- (en) Référence Animal Diversity Web : Echinodermata
- (en) Référence NCBI : Echinodermata Klein, 1734 (taxons inclus)
- (en) Référence Catalogue of Life : Echinodermata (consulté le )
- (en) Référence UICN : taxon Echinodermata (consulté le )
Liens externes[modifier | modifier le code]
- (en) Christopher L. Mah, « The Echinoblog », blog scientifique d'un spécialiste des échinodermes.
- « Les échinodermes », sur le site du Muséum national d'histoire naturelle.
- Antoine Morin, « Les Échinodermes », Université d'Ottawa.
- « Cours sur les échinodermes », sur usma.plongee.free.fr
- « Échinodermes », Site sur les échinodermes (dont les oursins) du MNHN et LIS.
- (en) Andreas Kroh, « The Echinoderm Files », sur www.marinespecies.org.
- (en) Julian Fell, Ben Wigham, Yannick Dewael, Alexander Kerr, Jean-Pierre Feral, « Echinoderm world records », sur Invertebrates.si.edu.
- (en) Christopher Mah, « Five Points About The Fossil History of Echinoderms », sur Echinoblog, .
- (en) HS Science Videos, « Shape of Life: Echinoderms - The Ultimate Animal », sur Youtube, .
- Ressources relatives au vivant :
- Animal Diversity Web
- Australian Faunal Directory
- BioLib
- Dyntaxa
- EPPO Global Database
- Paleobiology Database
- Global Biodiversity Information Facility
- iNaturalist
- Interim Register of Marine and Nonmarine Genera
- Nederlands Soortenregister
- New Zealand Organisms Register
- Système d'information taxonomique intégré
- World Register of Marine Species
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
Notes et références[modifier | modifier le code]
- http://www.marinespecies.org/aphia.php?p=taxdetails&id=1806 WoRMS taxon details
- Article « Échinodermes » dle l' Encyclopédie Larousse [1].
- (en) Christopher Mah, « Why five ? Mysteries in Symmetry! », sur Echinoblog, .
- Christopher Taylor, « Echinodermata », sur Palaeos (consulté le ). Nous ne retenons pas la classification proposée par ce site, car elle n'est pas consensuelle.
- Tree of Life Web Project, consulté le 2 mars 2014
- (en) « Echinodermata: Morphology », sur UCMP.Berkeley.edu (consulté le ).
- E. Tortonese & C. Vadon, « Oursins et holothuries (échinodermes) », sur www.fao.org.
- Alain Guille, Pierre Laboute et Jean-Louis Menou, Guide des étoiles de mer, oursins et autres échinodermes du lagon de Nouvelle-Calédonie, ORSTOM, , 244 p. (lire en ligne).
- Rich Mooi & Bruno David (2000) "What a new model of skeletal homologies tells us about asterid evolution" American Zoologist 40: 326-339.
- « Madréporite », sur Echinologia.
- (en) « Internal organs », sur l'Echinoid Directory du Natural History Museum.
- (en) Christopher Mah, « Starfish Feeding : When Starfish Capture moving Prey », sur Echinoblog, .
- (en) Formery, L., Peluso, P., Kohnle, I. et al., « Molecular evidence of anteroposterior patterning in adult echinoderms », Nature, vol. 623, , p. 555–561 (DOI 10.1038/s41586-023-06669-2).
- (en) Christopher Mah, « What are the Deepest known echinoderms ? », sur Echinoblog, .
- (en) Paola Rinelli, « A synthesis of the echinoderm faune of the Tyrrhenian sea », Rapp. Comm. int. Mer Médit., vol. 35, (lire en ligne).
- Chantal Conand, Sonia Ribes-Beaudemoulin, Florence Trentin, Thierry Mulochau et Émilie Boissin, Oursins, étoiles de mer & autres échinodermes : Biodiversité de La Réunion, La Réunion, Les éditions du Cyclone, , 168 p. (ISBN 979-10-94397-04-6).
- (en) FR Lillie, « The Fertilizing Power of Sperm Dilutions of Arbacia », Proc. Natl. Acad. Sci. U.S.A., vol. 1, no 3, , p. 156–60 (PMID 16575966, PMCID 1090763, DOI 10.1073/pnas.1.3.156)
- (en) MV Inamdar, T Kim, YK Chung, et al., « Assessment of sperm chemokinesis with exposure to jelly coats of sea urchin eggs and resact: a microfluidic experiment and numerical study. », J. Exp. Biol., vol. 210, no Pt 21, , p. 3805–20 (PMID 17951422, DOI 10.1242/jeb.005439, lire en ligne)
- (en) « Reproduction and life history », sur l'Echinoid Directory du Natural History Museum.
- (en) S. Ooka, M. Takeda, M. Komatsu et C. Conand, « Sexual reproduction of the small fissiparous seastar Aquilonastra conandae (Asteroidea: Asterinidae) in La Réunion Island », dans Larry G. Harris, S. Anne Böttger, Charles W. Walker et Michael P. Lesser, Echinoderms: Durham : Proceedings of the 12th International Echinoderm Conference, 7-11 August 2006, Durham, New Hampshire, U.S.A., Londres, Taylor & Francis, , 467-472 p. (ISBN 978-0-415-40819-6, DOI 10.1201/9780203869543-c72, lire en ligne).
- (en) Christopher Mah, « Deep-Sea Swimming Sea Cucumbers and the "most bizarre holothurian species in existence" ! », sur Echinoblog, .
- (en) Christopher Mah, « Some Stunning & Swimming Feather Star (Crinoid) Pics », sur Echinoblog, .
- Cohen-Rengifo, M., Moureaux, C., Dubois, P., & Flammang, P. (2017). Attachment capacity of the sea urchin Paracentrotus lividus in a range of seawater velocities in relation to test morphology and tube foot mechanical properties. Marine Biology, 164(4), 79|résumé
- (en) Christopher Mah, « Echinoderm Tube Feet Don't Suck! They Stick! », sur Echinoblog, .
- (en) Christopher Mah, « Holding on in a Rough World: Colobocentrotus atratus - the Shingle Urchin », sur Echinoblog,
- (en) Christopher L. Mah, « What (and How) do Sea Urchins Eat? Sea Urchin Feeding Roundup ! », sur The Echinoblog,
- (en) « Lifestyle - sand dollars », sur l'Echinoid Directory du Natural History Museum.
- (en) Christopher Mah, « Dermechinus horridus ! Trick or Treat ? », sur Echinoblog, (consulté le )
- (en) Christopher Mah, « What Eats Starfish ? (aka What do starfish fear ?) - The Top Starfish Predator : Solaster dawsoni », sur Echinoblog, .
- Philippe Bourjon, « Synapta maculata », sur Sous Les Mers (consulté le ).
- (en) Christopher Mah, « What Are the World's LARGEST Starfish ? », sur Echinoblog, .
- (en) « Thelenota anax », sur sealifebase.org.
- (en) Christopher Mah, « What we know about the world's most venomous sea urchin Toxopneustes fits in this blog post! », sur Echinoblog, (consulté le ).
- (en) Christopher Mah, « "The Most Common brittle star in the world" ? The tropical-global invasion of Ophiactis savignyi », sur Echinoblog, .
- Philippe Le Granché et Frédéric Ducarme, « Echinometra mathaei (Blainville, 1825) », sur DORIS, (MNHN), .
- (en) Christopher Mah, « Five Points About The Fossil History of Echinoderms », sur Echinoblog, .
- Christopher Taylor, « Homalozoa », sur Palaeos (consulté le )
- (en) M. Reich, « How many species of fossil holothurians are there? », Echinoderms in a Changing World, (ISBN 978-1-138-00010-0).
- Aristote, Histoire des Animaux, vol. IV, (lire en ligne).
- Pline l'Ancien, Histoire Naturelle, vol. IX, (lire en ligne).
- (en) A.V. Smirnov, « System of the Class Holothuroidea », Paleontological Journal, vol. 46, no 8, (DOI 10.1134/s0031030112080126, lire en ligne).
- Guillaume Rondelet, De piscibus marinis, libri XVIII, in quibus veræ piscium effigies expressæ sunt, Lyon, apud Matthiam Bonhomme, 1554.
- Daubenton, Jaucourt, L’Encyclopédie, 1re édition, 1751 (Tome 11, p. 716-718). lire en ligne.
- (en) John M. Lawrence, Sea Urchins : Biology and Ecology, Londres, Elsevier, (lire en ligne)
- (en) Christopher Mah, « The Big Echinoderm Big Book Blog », sur Echinoblog, .
- (en) « The Echinoderm Newsletter Archive », sur marinespecies.org/echinodermfiles.
- (en) Alexander Ziegler, Michel Jangoux, Georgy Mirantsev et Andreas Kroh, « Historical aspects of meetings, publication series, and digital resources dedicated to echinoderms », Zoosystematics and Evolution, vol. 90, no 1, , p. 45-56 (DOI 10.3897/zse.90.7201, lire en ligne).
- (en) Dr. Christopher Mah, « Brittle Star Diversity! How many are there and where do they live? », sur The Echinoblog, (consulté le ).
- World Register of Marine Species, consulté le 24 mars 2018
- « Observations > Echinodermes », sur iNaturalist, .
- World Register of Marine Species, consulté le 28 août 2013
- Integrated Taxonomic Information System (ITIS), www.itis.gov, CC0 https://doi.org/10.5066/F7KH0KBK, consulté le 28 août 2013
- NCBI, consulté le 28 août 2013
- BioLib, consulté le 5 janvier 2018
- M. J. Telford, C. J. Lowe, C. B. Cameron, O. Ortega-Martinez, J. Aronowicz, P. Oliveri et R. R. Copley, « Phylogenomic analysis of echinoderm class relationships supports Asterozoa », Proceedings of the Royal Society B: Biological Sciences, vol. 281, no 1786, , p. 20140479–20140479 (DOI 10.1098/rspb.2014.0479)
- « La Ballade des échinodermes », sur senscritique.com.
- Ferrario, C., Leggio, L., Leone, R., Di Benedetto, C., Guidetti, L., Coccè, V., ... & Sugni, M. (2017). Marine-derived collagen biomaterials from echinoderm connective tissues. Marine environmental research, 128, 46-57.