
Crinoidea

| Règne | Animalia |
|---|---|
| Sous-règne | Bilateria |
| Infra-règne | Deuterostomia |
| Embranchement | Echinodermata |
| Sous-embr. | Crinozoa |
Sous-classes de rang inférieur
- Articulata
- †Cladida (éteint)
- †Flexibilia (éteint)
- †Camerada (éteint)
- †Disparida (éteint)
Les Crinoïdes (Crinoidea) forment une classe d'animaux marins appartenant à l'embranchement des échinodermes.
Les crinoïdes sont des animaux sessiles (fixes) ou vagiles (mobiles) ressemblant à des plantes, mais pourvus d'un squelette calcaire articulé, d'une sorte de « racine » (éventuellement placée au bout d'une « tige »), et d'un « calice » muni de longs bras flexibles qui leur permettent de filtrer dans l'eau le plancton dont ils se nourrissent.
Étant des échinodermes, leurs plus proches parents dans le monde vivant sont les oursins, les étoiles de mer, les ophiures et les holothuries. Les premiers crinoïdes sont attestés à l'Ordovicien, période débutant il y a 500 millions d'années environ[1]. Malgré une morphologie rappelant celle d'un végétal — forme à laquelle ils doivent le nom commun de « Lys de mer » — les crinoïdes (du grec krinon, lys, et eidos, forme) sont des animaux (métazoaires)[2].
Description
[modifier | modifier le code]

Les crinoïdes possèdent un squelette calcaire dont les éléments, souvent très segmentés, sont organisés suivant une symétrie secondaire pentaradiaire (symétrie centrale d'ordre 5, en étoile), à l'instar des autres échinodermes (oursins, étoiles de mer…)[3]. Leur allure générale rappelle une fleur ou une chevelure, thèmes récurrents dans leurs nombreux noms vernaculaires[4].
À l'origine, les crinoïdes ressemblent morphologiquement à une sorte de grande fleur accrochée au fond marin : ils possèdent un analogue de « tige » plus ou moins longue (de quelques cm jusqu'à 10 m ancrée au fond par un crampon et terminée par ce qu'on appelle une « thèque », dont la partie inférieure est appelée « calice », et la face supérieure le « tegmen »[5] et d'où s'élancent de longs bras segmentés, articulés et barbulés pouvant ressembler à des plumes (ou à une chevelure) et dont les barbules sont appelées « pinnules »[6]. Certains crinoïdes sont accrochés au fond directement par leur calice, mais d'autres perdent cette attache à la maturité et la remplacent par des sortes de griffes mobiles (notamment les comatules) appelées « cirrhes », ou se tiennent directement sur leurs bras les plus dorsaux, et certains enfin peuvent avoir une vie plus mobile, et nager. On distingue ainsi ordinairement les « crinoïdes vrais » (fixés, munis ou non d'une tige) et les comatules, qui n'ont pas de tige et peuvent se déplacer et s'attacher où elles veulent grâce à leurs cirrhes ou à leurs bras (même si elles passent souvent la majorité de leur vie de manière plus ou moins sédentaire). En réalité, dans les deux cas la forme juvénile est attachée par une tige, porte 5 bras et une bouche ventrale : toutes les modifications propres à chaque espèce sont ultérieures, notamment la perte de la tige à la maturité[7]. De nos jours, la plupart des crinoïdes contemporains sont des comatules, mais on trouve encore des crinoïdes arborant une forme classique de « lys de mer » et attachés au fond par un pédoncule, notamment dans les grandes profondeurs[8]. Le crinoïde pédonculé vivant le moins profond est Metacrinus rotundus, qui peut se rencontrer dès 100 mètres de profondeur au Japon[1]. Attention cependant, la distinction entre comatules et crinoïdes fixes est paraphylétique : on trouve des crinoïdes ayant restauré l'usage d'une tige chez certains genres de l'ordre des Comatulida, et si certains crinoïdes fixes sont bien des survivants de familles mésozoïques, la plupart demeurent en réalité d'apparition « récente » (c'est-à-dire autour de la fin du Crétacé)[9].
C'est en déployant leurs bras en éventail que ces organismes filtrent l'eau de mer et se procurent leur nourriture (principalement du plancton), qu'ils acheminent jusque vers leur bouche grâce à de petits cils couvrant les bras, les pinnules[3]. Ces bras peuvent mesurer jusqu'à 50 cm de long chez le genre Alloeocomatella[7]. Les pinnules sont molles et sécrètent une glu collante : elles sont la seule partie du corps dépourvu de squelette calcifié, ce qui leur assure une bonne souplesse. Les cinq rayons brachiaux qui partent du centre sont divisés en différentes sections, les brachitaxis, bornés par des ossicules divisés, les axillaires. Les sections ainsi formées sont constituées d'un ensemble de brachiales nommées selon leur position le long du bras: secundibrachiales, tertiobrachiales, quartobrachiales, etc. suivant l'espèce et le nombre de bras[10].
Les segments (ou « articles ») qui constituent la tige (et les bras) de ces animaux, et que l'on nomme « entroques », sont circulaires ou pentagonaux et présentent un canal central[5] (isolés, ils ressemblent à de petits boulons) et cinq petits canaux périphériques. De structure stéréomique très minéralisée[1], ils constituent couramment l'élément le mieux conservé de l'animal lors de la fossilisation. La rupture de la tige ne semble pas létale à l'animal, qui survivra s'il parvient à se refixer ailleurs[3]. On considère qu'en moyenne 80 % de la biomasse des crinoïdes est calcifiée[5], ce qui en fait des animaux extrêmement durs et donc difficiles à attaquer pour d'éventuels prédateurs.
Le calice est une structure plus ou moins ovoïde, formé pour sa part de 2, 3 ou 5 rangées de plaques calcaires polygonales fusionnées (jamais 4 chez les espèces actuelles)[1]. Il contient les viscères et les organes vitaux (mais l'appareil reproducteur est situé au bout des bras), dont l'« organe cloisonné », situé à la base du calice et qui fait office de cœur. La respiration se fait en partie par l'anus, par contractions permanentes. La face supérieure du calice est appelée « tegmen »[5]. Elle est souple et contient la bouche au centre (parfois excentrée), où se rejoignent les 5 sillons ambulacraires, et l'anus, parfois placé au sommet d'une petite colonne, ainsi que des pores respiratoires[9]. En prolongement des sillons ambulacraires rayonnent les bras articulés, généralement au nombre de 5 ou 10, plus rarement 2, 4 ou 6. Ces bras peuvent être simples, branchus ou très ramifiés (les pinnules étant la ramification la plus fine). Des sillons ambulacraires partent de la bouche et parcourent les bras : ils portent de petits organes mous (proches des podia des autres échinodermes[5]) souvent couverts d'un mucus collant, qui font circuler la nourriture des bras jusqu'à la bouche[3].
Comme tous les échinodermes, les crinoïdes sont pourvus d'un « système aquifère » : il s'agit d'un système vasculaire dans lequel circule de l'eau de mer purifiée aspirée par les hydropores présents sur le tegmen[5], et qui assure des fonctions circulatoires, respiratoires, nutritives et motrices, et assure la turgescence des podia[1].
-
 Tige, thèque et bras d'un crinoïde « vrai » (de la famille des Isselicrinidae).
Tige, thèque et bras d'un crinoïde « vrai » (de la famille des Isselicrinidae). -
 Comatule de Bennett (Anneissia bennetti)
Comatule de Bennett (Anneissia bennetti) -
 Tegmen d'une Lamprometra palmata. La bouche est au centre des cinq sillons ambulacraires, et l'anus au sommet de la petite colonne.
Tegmen d'une Lamprometra palmata. La bouche est au centre des cinq sillons ambulacraires, et l'anus au sommet de la petite colonne. -

-
 Gros plan sur les pinnules d'une Tropiometra carinata (parasitée par Myzostoma fuscomaculatum).
Gros plan sur les pinnules d'une Tropiometra carinata (parasitée par Myzostoma fuscomaculatum).





Écologie et comportement
[modifier | modifier le code]Locomotion
[modifier | modifier le code]
Les crinoïdes sont des animaux benthiques : ils vivent posés sur le fond ou plus souvent attachées à un support, les crinoïdes à tige étant sessiles (même si la rupture de leur tige peut entraîner un déplacement et une relocation de l'animal). Les comatules sont vagiles, capables de marcher (ou parfois de nager[11]) grâce à leurs bras articulés sur de courtes distances, ce qui leur sert notamment à échapper aux prédateurs ou à chercher les meilleurs sites de nourrissage ; mais leurs mœurs demeurent fondamentalement sédentaires. Seule la larve est capable de nager sur de grandes distances : elle fait partie du plancton, et peut se laisser dériver pendant plusieurs semaines voire mois, ce qui permet une bonne dispersion des espèces[3].
Reproduction et croissance
[modifier | modifier le code]Les crinoïdes sont des animaux généralement sexués (présence d'individus mâles et femelles) même si certaines espèces peuvent être hermaphrodites[12]. Il ne semble pas exister de dimorphisme sexuel, et en dehors des organes reproducteurs (invisibles sans dissection hors période de reproduction), il est impossible de distinguer les individus mâles et femelles[13].
La reproduction est gonochorique, et mâles et femelles relâchent leurs gamètes en même temps grâce à un signal phéromonal (généralement à la saison chaude), en pleine eau, où les œufs vont se féconder et se développer. Les larves (appelées doliolaria) évoluent parmi le plancton pendant quelques semaines, puis se fixent à un support pour entamer leur métamorphose[10],[14]. Toutefois, chez certaines espèces les juvéniles se développent sur la mère avant de nager vers d'autres sites[12]. Comme chez tous les échinodermes, la larve est structurée selon une symétrie bilatérale, contrairement à l'adulte qui suit une symétrie pentaradiaire[1]. Elle ressemble à un tonnelet parcouru de zones ciliées : une touffe apicale et cinq bandes, dont une dépression vestibulaire ventrale.
Après la métamorphose, la post-larve s'accroche au substrat au moyen d'une tige (stade appelé cistide) puis la pointe arrondie se différencie en 5 bras (stade appelé Pentocrinus ou Pentacrinus). Les crinoïdes pédonculés conserveront la tige toute leur vie, alors que celle des comatules régressera jusqu'à disparaître à la maturité, remplacée par les cirrhes[1]. Le nombre de bras est initialement de 5, puis des divisions apparaissent chez la plupart des espèces, pour former parfois plusieurs centaines de bras.
-
 Stade larvaire d'une Antedon bifida
Stade larvaire d'une Antedon bifida -
 Stade juvénile (« pentacrinus ») d'une Antedon bifida
Stade juvénile (« pentacrinus ») d'une Antedon bifida -
 Jeune adulte d'Antedon bifida, libéré de sa tige.
Jeune adulte d'Antedon bifida, libéré de sa tige.



Alimentation
[modifier | modifier le code]
Les crinoïdes sont des animaux suspensivores. Leurs longs bras ramifiés munis de pinnules filtrent l'eau, et retiennent les particules en suspension (plancton, neige marine, particules organiques en suspension...) en s'enroulant, avant de transmettre le butin aux tentacules collants (« podia ») qui tapissent les sillons ambulacraires[10]. La nourriture circule ensuite le long de ces sillons comme sur un tapis roulant, pour arriver finalement à la bouche, d'où elle passera dans le tube digestif. Les déchets sont relâchés par l'anus lors des contractions respiratoires[14].
Les proies les plus communes des crinoïdes incluent des micro-algues comme des protistes (diatomées, dinoflagellées, foraminifères, tintinnides...), des larves d'invertébrés, de petits crustacés planctoniques (copépodes, ostracodes) et des particules détritiques[15]. L'analyse du contenu stomacal des crinoïdes montre que leur mode de capture est peu spécifique, et inclut également des particules de sable[15]. Il semble par ailleurs y avoir des variations significatives dans l'alimentation des crinoïdes en fonction de l'espèce, de l'environnement et de la saison, les espèces vivant plus profond étant moins sélectives et se nourrissant plus souvent de détritus[15].
Certaines espèces très particulières (notamment de la famille des Holopodidae, en forme de « poing ») pourraient cependant être capables de piéger des poissons ou d'autres animaux macroscopiques actifs[16].

Prédateurs
[modifier | modifier le code]
Les crinoïdes sont des animaux coriaces, au corps dur et grêle, et n'attirent donc que peu de prédateurs[5]. Seuls certains prédateurs très omnivores à la mâchoire solide en consomment occasionnellement. Parmi ces prédateurs on retrouve surtout de gros poissons (Balistidae, Tetraodontidae, Sparidae, les plus classiques étant Balistoides conspicillum et Chrysophrys auratus[17]), certains crabes (comme Oregonia gracilis[17]), étoiles de mer (Pycnopodia helianthoides[17]) ou, surtout en profondeur, des oursins[18].
La prédation par les oursins est observée dans le registre fossile dès le mésozoïque (et encore aujourd'hui dans les abysses), et il est possible que cette prédation ait joué un rôle dans la domination actuelle des comatules mobiles sur les crinoïdes fixes à tige[18].
Les bras des crinoïdes peuvent se couper de manière nette entre les articles, préservant ainsi le reste de l'animal lors d'une confrontation avec un prédateur (on parle d'autotomie, comme avec la queue des lézards). Comme souvent chez les échinodermes, les capacités de régénération sont ensuite impressionnantes, et les bras perdus sont rapidement reconstitués[17]. Même la masse viscérale peut être régénérée, en moins de 3 semaines chez l'espèce tropicale Himerometra robustipinna[17].

Symbioses et parasites
[modifier | modifier le code]
Les crinoïdes sont des organismes passifs, dont le corps dur attire peu les prédateurs : ils fournissent donc un refuge idéal à de nombreux petits animaux, qui se dissimulent entre leurs pinnules. La plupart de ces espèces adoptent une livrée striée, imitant les pinnules de l'hôte pour mieux s'y camoufler, mais certaines petites crevettes préfèrent être simplement transparentes.
Parmi les hôtes fréquents des crinoïdes, on compte notamment plusieurs espèces de crevettes nettoyeuses de la famille des Palaemonidae (notamment des genres Laomenes, Periclimenes ou Manipontonia, comme L. amboinensis, L. cornutus, P. pilipes ou P. ceratophthalmus), dont la couleur mime généralement celle de l'hôte. On trouve parfois aussi des crevettes plus grosses, comme la crevette-pistolet Synalpheus demani. Les Galathées des crinoïdes (genre Allogalathea) vivent également en symbiose avec les crinoïdes, qu'elles débarrassent des parasites en échange d'un abri entre leurs bras. Le petit crabe Tiaramedon spinosum est fréquent chez les petites comatules tropicales de faible profondeur de l'Indo-Pacifique, et l'encore plus petit Harrovia albolineata se rencontre plutôt dans le Pacifique (ainsi que, moins fréquemment, Harrovia elegans et Ceratocarcinus longimanus)[19]. Certains poissons du genre Discotrema s'y retrouvent également, notamment les juvéniles, ainsi que le poisson-fantôme arlequin, dont le corps frangé imite les pinnules des comatules. Dans le Pacifique, on peut aussi y trouer le calmar pygmée des crinoïdes, Idiosepius pygmaeus[20]. Plusieurs vers polynoides leur sont aussi associés, sans qu'il soit facile de discerner le bénéfice ou coût pour l'hôte.
Les crinoïdes peuvent aussi être victimes de parasites, notamment de la part de vers marins comme Myzostoma fuscomaculatum[21].
-

-

-

-

-

-
 Poisson-fantôme arlequin (Solenostomus paradoxus)
Poisson-fantôme arlequin (Solenostomus paradoxus) -
 Myzostoma fuscomaculatum (parasite)
Myzostoma fuscomaculatum (parasite)
.jpg)
.jpg)



_1.png)

Habitat et répartition
[modifier | modifier le code]
Comme tous les échinodermes, les crinoïdes sont des animaux strictement marins. On ne connaît pas à ce jour de fossile de crinoïde en milieu fluvial ou lacustre, mais leur répartition en profondeur est très étendue. Les espèces actuelles sont principalement cantonnées aux mers tropicales (ils sont très abondants aux Philippines et en Indonésie), cependant il en existe des espèces d'eaux tempérées voire froides : par exemple deux espèces en Méditerranée (l’Antedon mediterranea et le Leptometra phalangium) ainsi qu'une espèce dans l'Atlantique nord (l’Antedon bifida, cette dernière visible depuis le Portugal jusqu'en Norvège). Les seules grandes mers ouvertes dépourvues de crinoïdes semblent être la mer Noire (qui ne comporte quasiment aucun échinoderme) et la mer Baltique[1].
De nombreuses espèces habitent également les grandes profondeurs, où elles se nourrissent de plancton et de neige marine. La plupart des crinoïdes à tiges ayant subsisté jusqu'à notre époque vivent ainsi à plusieurs centaines de mètres de profondeur (on en trouve par exemple au large du golfe de Gascogne)[3]. Plusieurs genres sont même inféodés aux abysses extrêmes (« zone hadale »), notamment dans la famille des Bathycrinidae (crinoïdes à tige), dont des individus ont été filmés entre 8 175 et 9 050 m de profondeur dans les fosses océaniques du Pacifique nord[22]. Cependant, la majorité des espèces fixes vivent plutôt dans la zone bathyale, c'est-à-dire entre 200 et 1 000 m de profondeur[9].
De manière générale, à faible profondeur on ne trouve que des comatules (donc des crinoïdes mobiles pourvus de cirrhes plutôt que d'une tige), alors qu'à plus grande profondeur on trouve aussi bien des crinoïdes fixes que mobiles. L'espèce à tige vivant le moins profond semble être Metacrinus rotundus, que l'on peut trouver au Japon dès 100 m de profondeur[9]. Malgré la grande diversité des crinoïdes, les espèces rencontrées à faible profondeur ne sont réparties que sur 7 familles, comportant au total 370 espèces (dont beaucoup d'abyssales ou mésophotiques).
L'habitat de prédilection des crinoïdes est les récifs de corail, riches en anfractuosités dans lesquelles se cacher et en reliefs sur lesquels s'accrocher pour capter le courant. Cependant, dans certaines mers où la nourriture est abondante, on peut observer de vastes populations de crinoïdes sur tous types de terrains, y compris des sols meubles[23].
-

-

-
 Stephanometra indica est une des espèces les plus communes de l'Indo-Pacifique.
Stephanometra indica est une des espèces les plus communes de l'Indo-Pacifique. -
 Leptometra celtica, une espèce européenne des eaux froides de l'Atlantique nord.
Leptometra celtica, une espèce européenne des eaux froides de l'Atlantique nord. -
 Comatule Ptilometra australis en Australie du sud.
Comatule Ptilometra australis en Australie du sud. -
 En abondance dans une faille en Afrique du Sud.
En abondance dans une faille en Afrique du Sud. -
 Un crinoïde et une comatule à grande profondeur.
Un crinoïde et une comatule à grande profondeur.

.jpg)





Classification
[modifier | modifier le code]




Les bases de la classification moderne des crinoïdes ont été posées dans la Monograph of the Existing Crinoidea d'Austin H. Clark, publiée à partir de 1915[24], sur la base de critères essentiellement squelettiques. Cependant, les données génétiques récentes suggèrent que certains critères squelettiques utilisés dans la classification du XXe siècle sont en réalité trompeurs, et la taxinomie des crinoïdes est actuellement en pleine révision, notamment les comatules depuis 2014 par l'équipe américaine de Charles G. Messing[25],[7].
On compte à l'heure actuelle environ 650 espèces de crinoïdes (dont moins d'une centaine de pédonculés[1]), ce qui en fait le groupe le plus réduit de tous les échinodermes, et le moins diversifié avec seulement 142 genres classés en 32 familles actuelles, réparties en 4 ordres[26]. Seules 55 espèces actuelles n'appartiennent pas à l'ordre des Comatulida.
|
Selon World Register of Marine Species (8 avril 2015)[27] : ___
|
Selon ITIS (2 décembre 2013)[28] et Animal Diversity Web (2 décembre 2013)[29] :
Selon NCBI (2 décembre 2013)[30] :
|
- Galerie représentatives des ordres actuels, et de quelques-uns des principaux ordres fossiles
-

-

-

-

-

-

-

-

-
 Crinometra brevipinna (individus pâles), une Charitometridae
Crinometra brevipinna (individus pâles), une Charitometridae -
-

-
 Une Bathycrinidae (espèce abyssale qui a restauré l'usage d'une tige)
Une Bathycrinidae (espèce abyssale qui a restauré l'usage d'une tige) -
 Guillecrinus neocaledonicus, une Guillecrinidae (idem)
Guillecrinus neocaledonicus, une Guillecrinidae (idem) -
 Une Phrynocrinidae (idem)
Une Phrynocrinidae (idem) -
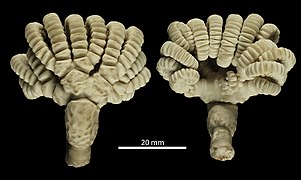 Neogymnocrinus richeri, l'unique Sclerocrinidae
Neogymnocrinus richeri, l'unique Sclerocrinidae -
 Holopus sp., une Holopodidae
Holopus sp., une Holopodidae -
 Fossile d'Encrinus liliiformis, un Encrinida
Fossile d'Encrinus liliiformis, un Encrinida -
 Hyocrinus sp., une Hyocrinida
Hyocrinus sp., une Hyocrinida -
 Colonie de Neocrinus decorus, une Isocrinidae
Colonie de Neocrinus decorus, une Isocrinidae -

-

.jpg)






_002.jpeg)












Origine et évolution
[modifier | modifier le code]La structure extrêmement calcifiée du corps des crinoïdes leur assure une excellente conservation et fossilisation après leur mort : ainsi, ces animaux sont relativement bien connus des paléontologues, et constituent un groupe-repère d'importance capitale[5]. Plus souvent encore que les crinoïdes entiers, on trouve dans le registre fossile des entroques[31], segments de leur tige ou de leurs bras, dispersés après la mort de l'animal et fossilisés séparément, souvent dans des calcaires dits « à entroques » ou « à encrines » ; on en trouve aussi dans certains marbres où les cercles blancs des entroques se repèrent aisément[32].
Les ancêtres des crinoïdes apparaissent au Cambrien[33] entre -541,0 ± 1,0 Ma et -485,4 ± 1,9 Ma : il s'agit d'un groupe comportant également les cystoïdes, les blastoïdes et les éocrinoïdes (tous éteints). Les crinoïdes proprement-dits sont bien identifiés vers -485,4 Ma, à l'Ordovicien[33], avec un pic de diversité au Mississippien[9] soit de -358 à -323 Ma. Ils sont les seuls de leur groupe à survivre à l'extinction Permien-Trias (il y a environ 252 millions d'années), ce qui leur permet une nouvelle radiation évolutive à partir du Mésozoïque, sans pour autant retrouver leur diversité du Paléozoïque : au fil des phases d'extinction qui suivirent, la plupart des formes qui ont survécu jusqu'à nos jours sont des espèces mobiles comme les comatules[9]. Les crinoïdes à tige sont aujourd'hui pour la plupart cantonnées aux abysses[34].
Pendant le Paléozoïque et le Mésozoïque, les crinoïdes sessiles abondaient et constituaient de véritables prairies sous-marines. Certains groupes disparus avaient développé des adaptations particulières qui n'existent plus aujourd'hui : au Dévonien, les Ammonicrinus vivant dans la zone tidale pouvaient s'enrouler sur eux-mêmes à la manière des cloportes pour se protéger. Les Crotalocrinidae formaient des colonies construisant de petits récifs. Au Mésozoïque, les Cyrtocrinides Uintacrinidés semblaient pélagiques, équipés d'un flotteur à la place de la tige[14].
Les comatules semblent être apparues seulement entre le Trias[9] et le Jurassique[1], peut-être sous la pression des prédateurs des crinoïdes sessiles, tels les oursins[18]. Cependant, il faut attendre la fin du Miocène pour que les comatules supplantent leurs cousins fixes[9].
-
 Fossile d'Agaricocrinus americanus (carbonifère)
Fossile d'Agaricocrinus americanus (carbonifère) -
 Crinoïdes fossiles exceptionnellement bien préservés.
Crinoïdes fossiles exceptionnellement bien préservés. -
 Fossile de thèque d'Actinocrinus.
Fossile de thèque d'Actinocrinus. -

-
 Crinoïdes du début du Carbonifère.
Crinoïdes du début du Carbonifère. -

-
 Crampons de crinoïdes de l'ordovicien.
Crampons de crinoïdes de l'ordovicien. -

-










Selon BioLib (27 décembre 2017)[35] :
- sous-classe Articulata Simms, 1988
- ordre Comatulida
- ordre Hyocrinida Rasmussen, 1978
- ordre Isocrinida
- ordre Uintacrinida A. H. Clark, 1908 †
- sous-classe Camerata Wachsmuth & Springer, 1885 †
- ordre Monobathrida Moore & Laudon, 1943 †
- sous-classe Cladida Moore & Laudon, 1943 †
- ordre Cyathocrinida Bather, 1899 †
- ordre Dendrocrinida Bather, 1899 †
- famille Codiacrinidae Bather, 1890 †
- sous-classe Disparida Moore & Laudon, 1943 †
- ordre Pisocrinida Ausich & Copper, 2010 †
- famille Acolocrinidae C. E. Brett, 1980 †
- famille Allagecrinidae Carpenter & Etheridge, 1881 †
- famille Anamesocrinidae Goldring, 1923 †
- famille Anomalocrinidae Wachsmuth & Springer, 1886 †
- famille Apodasmocrinidae T. J. Frest, H. L. Strimple & M. R. McGinnis, 1979 †
- famille Belemnocrinidae S.A. Miller, 1883 †
- famille Calceocrinidae Meek & Worthen, 1869 †
- famille Catillocrinidae Wachsmuth & Springer, 1886 †
- famille Haplocrinitidae Bassler, 1938 †
- famille Heterocrinidae Zittel, 1879 †
- famille Holynocrinidae Bouška, 1948 †
- famille Homocrinidae Kirk, 1914 †
- famille Iocrinidae Moore & Laudon, 1943 †
- famille Myelodactylidae Miller, 1883 †
- famille Pygmaeocrinidae Strimple, 1963 †
- famille Ramacrinidae Prokop, 1977 †
- famille Zophocrinidae Miller, 1892 †
- sous-classe Flexibilia von Zittel, 1895 †
- ordre Sagenocrinida Springer, 1913 †
- ordre Taxocrinida Springer, 1913 †
- famille Edriocrinidae S. A. Miller, 1889 †
- infra-classe Cyclici Moore & Jeffords, 1968 †
- famille Mediocrinidae Stukalina, 1964 †
- famille Salairocrinidae Dubatolova, 1971 †
- infra-classe Pentameri Moore & Jeffords, 1968 †
- ordre Angulata Stukalina, 1967 †
- famille Anthinocrinidae Yeltyschewa & Sisova in Schewtschenko, 1966 †
- famille Pentacauliscidae Moore & Jeffords, 1968 †
- ordre Diplobathrida †
- ordre Encrinida †
- ordre Millericrinida Sieverts-Doreck, 1952 †
- famille Asperocrinidae Stukalina, 1975
- famille Cyclomischidae Moore & Jeffords, 1968 †
Dans la culture
[modifier | modifier le code]Les crinoïdes sont des animaux discrets et facilement mystérieux pour les baigneurs, et demeurent donc relativement mal connus du grand public. Ils figurent cependant dans le célèbre roman sous-marin de Jules Verne, Vingt mille lieues sous les mers, décrivant non sans exagération « des comatules ambulantes, larges d’un mètre, et dont la pourpre rougissait les eaux », et plus loin (et plus profond) « Alors la vie animale n’était plus représentée que par des encrines, des étoiles de mer, de charmantes pentacrines tête de méduse, dont la tige droite supportait un petit calice ».
Au Japon, où les invertébrés ont une place plus importante dans le folklore[36], on note deux pokémons dessinés d'après ces animaux dans le jeu vidéo éponyme : Lilia, et son évolution Vacilys[37].
Galerie
[modifier | modifier le code]-
 Vue d'un crinoïde.
Vue d'un crinoïde. -
 Crinoïde fixe de grande profondeur. Les entroques sont bien visibles.
Crinoïde fixe de grande profondeur. Les entroques sont bien visibles. -
 Autre crinoïde « vrai » à Hawaii.
Autre crinoïde « vrai » à Hawaii. -

-

-

-
 Une crevette symbiotique (Synalpheus demani) cachée sur le tegmen d'un crinoïde.
Une crevette symbiotique (Synalpheus demani) cachée sur le tegmen d'un crinoïde. -

-
 Détails d'une comatule.
Détails d'une comatule. -

-
 Comatula purpurea vue du dessous.
Comatula purpurea vue du dessous. -
 Fossile dans la collection du Musée des enfants d'Indianapolis.
Fossile dans la collection du Musée des enfants d'Indianapolis. -
 Florometra serratissima en grandes profondeurs.
Florometra serratissima en grandes profondeurs. -
 Assemblage multicolore à Bali.
Assemblage multicolore à Bali.
.jpg)








.jpg)
_underside.jpg)



Voir aussi
[modifier | modifier le code]- Échinoderme
- Echinodermata (classification phylogénétique)
- Comatule
- Espèce panchronique
- Echinodermes de Méditerranée
- cystoïdes, blastoïdes et éocrinoïdes
- Entroque
Références taxinomiques
[modifier | modifier le code]- (en) Référence WoRMS : Crinoidea Miller, 1821 (+ liste ordres + liste familles)
- (en) Référence Paleobiology Database : Crinoidea Miller 1821
- (fr + en) Référence ITIS : Crinoidea Miller, 1821
- (en) Référence Tree of Life Web Project : Crinoidea
- (en) Référence Animal Diversity Web : Crinoidea
- (en) Référence Catalogue of Life : Crinoidea (consulté le )
- (en) Référence NCBI : Crinoidea (taxons inclus)
Bibliographie
[modifier | modifier le code]- (en) Michel Roux, Charles G. Messing et Nadia Améziane, « Artificial keys to the genera of living stalked crinoids (Echinodermata) », Bulletin Of Marine Science, vol. 70, no 3, , p. 799–830 (lire en ligne).
- Chantal Conand, Sonia Ribes-Beaudemoulin, Florence Trentin, Thierry Mulochau et Émilie Boissin, Oursins, étoiles de mer & autres échinodermes : Biodiversité de La Réunion, La Réunion, Les éditions du Cyclone, , 168 p. (ISBN 979-10-94397-04-6).
- Alain Guille, Pierre Laboute et Jean-Louis Menou, Guide des étoiles de mer, oursins et autres échinodermes du lagon de Nouvelle-Calédonie, ORSTOM, , 244 p. (lire en ligne)
- René Koehler, Faune de France : 1 : Échinodermes, Paris, Librairie de la Faculté des Sciences, (lire en ligne).
- (en) A.M. Clark et F.W.E. Rowe, Monograph of Shallow-water Indo-West Pacific Echinoderms, Londres, Trustees of the British Museum (Natural History), , 238 p. (lire en ligne).
- (en) Maria Byrne et Timothy O'Hara, Australian echinoderms : Biology, Ecology and Evolution, CSIRO Publishing, (ISBN 978-1-4863-0763-0, lire en ligne).
- Frédéric Ducarme, Étoiles de mer, oursins et autres échinodermes de Mayotte et sa région, Les Naturalistes de Mayotte, , 336p (ISBN 978-2-9521543-5-2).
Liens externes
[modifier | modifier le code]- (en) Charles Messing, « Charles Messing's Crinoids page », sur www.nova.edu.
- (en) [vidéo] « Vidéo d'introduction aux crinoïdes par Charles Messing », sur YouTube
- (en) Charles Messing, « Sea Star on a Stick: Introducing Crinoids », sur Vimeo, .
- (en) « Guide d'identification des crinoïdes australiens », sur australianmuseum.net.au.
- Denis Tournassat, « Crinoïdes : Lys de mer et comatules », sur cap-recifal.com, .
- « Crinoïdes », sur Échinodermes.org, site sur les échinodermes du MNHN et LIS.
- « Crinoïdes et comatules », sur DORIS, encyclopédie marine en ligne de la FFESSM.
- François Cornu, « Crinoïdes et comatules », sur Sous Les Mers.
- (en) Christopher Mah, « The Echinoblog », site du Dr Christopher Mah, zoologue spécialiste des échinodermes.
- Ressources relatives au vivant :
Notes et références
[modifier | modifier le code]- (en) Charles Messing, « Introduction », sur Charles Messing's Crinoids page.
- www.jeanlouisetienne.com Les Comatules (Crinoïdes) (consulté le 1er décembre 2014)
- « Les Crinoïdes », sur Cosmovisions.com (consulté le ).
- (en) Christopher Mah, « Crinoid Close ups! », sur Echinoblog, .
- (en) Charles Messing, « Introduction to morphology », sur Charles Messing's Crinoids page.
- Pinnula signifie « petite plume » en latin tardif.
- (en) Mindi M. Summers, Charles G. Messing et Greg W. Rouse, « The genera and species of Comatulidae (Comatulida: Crinoidea): taxonomic revisions and a molecular and morphological guide », Zootaxa, vol. 4268, no 2, , p. 151–190 (DOI 10.11646/zootaxa.4268.2.1, lire en ligne).
- (en) Christopher Mah, « Stalked Crinoid Round-Up », sur Echinoblog, .
- (en) Michel Roux, Charles Messing et Nadia Améziane, « Artificial keys to the genera of living stalked crinoids (Echinodermata) », Bulletin of Marine Science, vol. 70, no 3, , p. 799-830 (lire en ligne).
- Alain Guille, Pierre Laboute et Jean-Louis Menou, Guide des étoiles de mer, oursins et autres échinodermes du lagon de Nouvelle-Calédonie, ORSTOM, , 244 p. (lire en ligne).
- (en) Christopher Mah, « Some Stunning & Swimming Feather Star (Crinoid) Pics », sur Echinoblog, .
- (en) Obuchi, M., Fujita, Y., Nakano, Y. et al., « Reproductive biology and early life history of the hermaphroditic feather star Dorometra sesokonis (Echinodermata: Crinoidea) », Marine Biology, vol. 157, , p. 1191–1201 (DOI 10.1007/s00227-010-1399-3).
- (en) KH Chang, CP Chen, JC Chen, « Reproductive biology of the crinoid Comanthus parvicirrus and Comatella maculata (Echinodermata: Crinoidea) », Bull. Inst. zool. Acad. Sin, vol. 29, , p. 165-172.
- « Les crinoïdes », sur Fossile.fr (consulté le ).
- (en) Charles Messing, « Crinoid diet », sur www.nova.edu.
- (en) Christopher Mah, « Crinoid Crazy : Let's Meet Holopus », sur Echinoblog, .
- (en) Charles Messing, « Predation on living crinoids », sur www.nova.edu.
- (en) Christopher Mah, « Echinoderm Big Battle : Sea Urchins Feeding on Stalked Crinoids », sur Echinoblog, .
- Steven Weinberg, Découvrir la vie sous-marine : Mer Rouge océan Indien océan Pacifique, t. 1, Gap, , 560 p. (lire en ligne).
- (en) Albert Kang, « Two-toned Pygmy Squid - Idiosepius pygmaeus », sur jungledragon.com.
- (en) Christopher Mah, « Fossil Parasites & Commensals : 5 Examples », sur Echinoblog, .
- (en) Christopher Mah, « What are the Deepest known echinoderms ? », sur Echinoblog, .
- (en) TS Tay et KS Tan, « Crinoid diversity in the subtidal non-coral reef habitats of Singapore », 15th International Echinoderm Conference, .
- (en) Christopher Mah, « The Big Echinoderm Big Book Blog », sur Echinoblog, .
- (en) M. M. Summers, C. G. Messing et G. W. Rouse, « Phylogeny of Comatulidae (Echinodermata: Crinoidea: Comatulida): A new classification and an assessment of morphological characters for crinoid taxonomy », Molecular Phylogenetics and Evolution, vol. 80, , p. 319-339 (DOI 10.1016/j.ympev.2014.06.030).
- World Register of Marine Species, consulté le 2 décembre 2013
- World Register of Marine Species, consulté le 8 avril 2015
- Integrated Taxonomic Information System (ITIS), www.itis.gov, CC0 https://doi.org/10.5066/F7KH0KBK, consulté le 2 décembre 2013
- Myers, P., R. Espinosa, C. S. Parr, T. Jones, G. S. Hammond, and T. A. Dewey. The Animal Diversity Web (online). Accessed at https://animaldiversity.org, consulté le 2 décembre 2013
- NCBI, consulté le 2 décembre 2013
- Définition du mot « entroque » [1].
- « Définition : calcaire à entroques », sur La Revue du Vin de France (consulté le ).
- Fossilworks Paleobiology Database, consulté le 13 janvier 2014
- (en) Christopher Mah, « When Sea Urchins attack : Crinoid evasion = Crinoid evolution », sur Echinoblog, .
- BioLib, consulté le 27 décembre 2017
- Frédéric Ducarme, « Pourquoi étudier les invertébrés ? Quelques arguments d’Aristote », sur No Bones, Smithsonian Institute, .
- (en) « On the Origin of Species: Lileep and Cradily », sur bulbanews.bulbagarden.net.