
Hémocyanine


L'hémocyanine, couramment symbolisée par Hc, est une métalloprotéine contenant du cuivre ayant pour fonction de transporter l'oxygène chez certains animaux parmi lesquels des arthropodes (crustacés, arachnides, certains insectes) et des mollusques (bivalves, gastéropodes, coléoïdes notamment). Elle contient deux cations cuivreux Cu+ qui se lient à une molécule O2 de manière réversible en devenant cuivriques Cu2+. L'oxygénation provoque un changement de couleur : la forme désoxygénée (à cations Cu+) est incolore, tandis que la forme oxygénée (à cations Cu2+) est bleue[1]. Contrairement à l'hémoglobine des érythrocytes des vertébrés, l'hémocyanine flotte librement dans l'hémolymphe sans être contenue dans des cellules sanguines. Les hémocyanines sont les deuxièmes transporteurs d'oxygène les plus utilisés parmi les êtres vivants après les hémoglobines.
Superfamille de l'hémocyanine
La superfamille de l'hémocyanine des arthropodes est composée des phénol oxydases, des hexamérines, des pseudohémocyanines ou cryptocyanines, et des récepteurs d'hexamérines.
- Les phénol oxydases sont des tyrosinases contenant du cuivre. Elles interviennent dans les processus de sclérotisation de la cuticule des arthropodes, dans la cicatrisation des plaies et dans le système immunitaire. Elles sont produites sous forme de proenzymes qui sont activés par clivage du peptide N-terminal.
- Les hexamérines sont des protéines de stockage courantes chez les insectes. Elles sont produites le corps gras des larves et sont associées aux mues et aux conditions nutritionnelles.
- Les séquences des pseudohémocyanines et cryptocyanines sont étroitement apparentées à celle de l'hémocyanine des crustacés. Elles possèdent une structure et une fonction semblables, mais sont dépourvues de sites de liaison au cuivre.
Les changements observés dans la phylogénie de la superfamille de l'hémocyanine sont étroitement liés à l'émergence de ces différentes protéines à travers les différentes espèces. La compréhension de ces changements passe par l'étude approfondie des changements de l'hémocyanine chez les arthropodes[2].



Structure et fonctionnement
Bien que la fonction respiratoire de l'hémocyanine soit semblable à celle de l'hémoglobine, sa structure moléculaire et son mécanisme de liaison à l'oxygène O2 en diffèrent sensiblement. Ainsi, là où l'hémoglobine utilise un groupe prosthétique héminique contenant un cation de fer, l'hémocyanine utilise une paire de cations de cuivre directement coordonnés chacun à trois résidus d'histidine de son site actif[3].
-

Désoxyhémocyanine. -
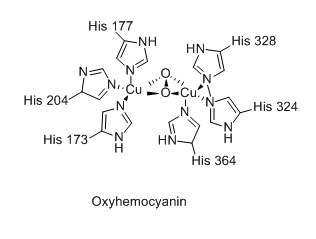
Oxyhémocyanine.


L'affinité de l'hémocyanine pour l'oxygène est supérieure à celle de l'hémoglobine, mais son efficacité globale comme transporteur d'oxygène est, dans la plupart des cas, plus faible que celle de l'hémoglobine. Elle est cependant supérieure dans les environnements froids[4]. On relève ainsi, parmi les organismes qui utilisent l'hémocyanine comme transporteur d'oxygène, des crustacés vivant dans des eaux froides à faible pression partielle d'oxygène ; il existe cependant également des arthropodes terrestres vivant dans des environnements chauds et oxygénés et qui utilisent malgré tout l'hémocyanine, comme les scorpions et les araignées[5].
L'hémocyanine est constituée de plusieurs sous-unités qui contiennent chacune une paire de cations de cuivre susceptible de se lier à une molécule d'oxygène. Chacune de ces sous-unités a une masse d'environ 75 kDa. Elles sont généralement homogènes, ou hétérogènes avec deux types de sous-unités. Elles s'organisent en dimères ou en hexamères selon les espèces, ces dimères et hexamères étant à leur organisés en chaînes ou en grappes dont la masse peut dépasser 1 500 kDa. En raison de la très grande taille de ces complexes, l'hémocyanine est généralement à l'état libre dans l'hémolymphe et non contenue dans des cellules sanguines, contrairement à l'hémoglobine[6].
Les hexamères sont caractéristiques de l'hémocyanine des arthropodes. Ainsi, celle de la tarentule Eurypelma californicum comprend quatre hexamères, soit 24 sous-unités, tandis que celle de la scutigère véloce, un chilopode, comprend six hexamères, soit 36 sous-unités, et celle des limules comprend huit hexamères 48 sous-unités. On trouve des hexamères simples chez Panulirus interruptus, une langouste, et chez le bathynome géant. Les sous-unités d'hémocyanine de crustacés contiennent environ 660 résidus d'acides aminés, tandis que celles des chélicérates en contiennent 625.
La plupart des hémocyanines se lient à l'oxygène de manière non-coopérative et sont capables de transporter environ le quart de la quantité d'oxygène transportée par l'hémoglobine. On observe cependant un effet coopératif chez certaines espèces d'arthropodes et chez les limules, avec un coefficient de Hill (en) compris entre 1,6[7],[8] chez Tachypleus gigas (une limule du Japon) et 3,0[9] chez Androctonus australis (un scorpion du Maghreb), valeurs qui dépendent également des conditions expérimentales ; à titre de comparaison, le coefficient de Hill de l'hémoglobine est généralement voisin de 3,0. L'hémocyanine présente dans ce cas une structure hexamérique, chaque sous-unité possédant un site de liaison à l'oxygène : la liaison d'une molécule d'oxygène à l'une de ces sous-unités accroît l'affinité des sous-unités voisines pour l'oxygène, selon un mécanisme allostérique. Ces études ont montré que les hexamères s'associent en complexes protéiques plus grands comprenant des douzaines d'hexamères. L'une de ces études suggère que l'effet coopératif est dépendant de ces arrangements de grande taille, de sorte que l'effet coopératif se propagerait entre hexamères[10]. Les profils de liaison de l'oxygène à l'hémocyanine sont affectés par la force ionique de la solution, et par son pH[11].
Activité catalytique
| N° EC | EC |
|---|---|
| N° CAS | |
| Cofacteur(s) | Cuivre |
| IUBMB | Entrée IUBMB |
|---|---|
| IntEnz | Vue IntEnz |
| BRENDA | Entrée BRENDA |
| KEGG | Entrée KEGG |
| MetaCyc | Voie métabolique |
| PRIAM | Profil |
| PDB | RCSB PDB PDBe PDBj PDBsum |
| GO | AmiGO / EGO |
L'hémocyanine est homologue des phénol oxydases (comme la tyrosinase) dans la mesure où elle possède un site actif semblable, utilisant une paire de cations de cuivre comme groupe prosthétique[12]. Dans tous les cas, une proenzyme inactive telle que l'hémocyanine, la tyrosinase et la catéchol oxydase (en) doit être préalablement activée par clivage du résidu d'acide aminé qui bloque l'entrée du canal conduisant au site actif. Les différences conformationnelles déterminent le type d'activité catalytique susceptible d'être réalisée par l'hémocyanine[13]. L'hémocyanine possède également une activité phénol oxydase, mais cependant plus lente ; une dénaturation partielle a pour effet de favoriser cette activité enzymatique en dégageant l'accès au site actif[1],[12].
Effet anticancéreux
L'hémocyanine de Concholepas concholepas (en) présente un effet thérapeuthique contre le cancer de la vessie chez la souris, ce qui en fait un objet de recherche pour d'éventuels traitements du cancer superficiel de la vessie.
L'hémocyanine de Megathura crenulata, dite KLH, est un stimulant immunitaire dérivé de glycoprotéines de ce mollusque. Lorsqu'elle est administrée in vitro, elle présente une activité significative contre la prolifération des cellules de cancer du sein, de cancer du pancréas et de cancer de la prostate. Une étude de 2003 a également suscité un intérêt concernant l'effet de l'hémocyanine de Megathura crenulata contre l'adénocarcinome de l'endobrachyœsophage[14].
Notes et références
- (en) Christopher J. Coates et Jacqueline Nairn, « Diverse immune functions of hemocyanins », Developmental & Comparative Immunology, vol. 45, no 1, , p. 43-55 (PMID 24486681, DOI 10.1016/j.dci.2014.01.021, lire en ligne)
- (en) Thorsten Burmester, « Molecular Evolution of the Arthropod Hemocyanin Superfamily », Molecular Biology and Evolution, vol. 18, no 2, , p. 184-195 (PMID 11158377, DOI 10.1093/oxfordjournals.molbev.a003792, lire en ligne)
- (en) N. S. Rannulu et M. T. Rodgers, « Solvation of copper ions by imidazole: Structures and sequential binding energies of Cu+(imidazole)x, x = 1–4. Competition between ion solvation and hydrogen bonding », Physical Chemistry Chemical Physics, vol. 7, no 5, , p. 1014-1025 (PMID 19791394, DOI 10.1039/B418141G, Bibcode 2005PCCP....7.1014R, lire en ligne)
- (en) Michael Oellermann, Bernhard Lieb, Hans-O Pörtner, Jayson M. Semmens et Felix C. Mark, « Blue blood on ice: modulated blood oxygen transport facilitates cold compensation and eurythermy in an Antarctic octopod », Frontiers in Zoology (en), vol. 12, , p. 6 (PMID 25897316, PMCID 4403823, DOI 10.1186/s12983-015-0097-x, lire en ligne)
- (en) Anneli Strobel, Marian Y. A. Hu, Magalena A. Gutowska, Bernhard Lieb, Magnus Lucassen, Frank Melzner, Hans O. Pörtner et Felix C. Mark, « Influence of Temperature, Hypercapnia, and Development on the Relative Expression of Different Hemocyanin Isoforms in the Common Cuttlefish Sepia officinalis », Journal of Experimental Zoology. Part A, Ecological Genetics and Physiology, vol. 317, no 8, décembre, p. 2012 (PMID 22791630, DOI 10.1002/jez.1743, lire en ligne)
- (en) L. Waxman, « The structure of arthropod and mollusc hemocyanins », Journal of Biological Chemistry, vol. 250, no 10, , p. 3796-3806 (PMID 1126935, lire en ligne)
- (en) Nobuo Makino, « Hemocyanin from Tachypleus gigas. I. Oxygen-Binding Properties », Journal of Biochemistry, vol. 106, no 3, , p. 418-422 (PMID 2606894, lire en ligne)
- (en) Nobuo Makino, « Hemocyanin from Tachypleus gigas. II. Cooperative Interactions of the Subunits », Journal of Biochemistry, vol. 106, no 3, , p. 423-429 (PMID 2606895, lire en ligne)
- (en) Felix de Haas et Ernst F.J. van Bruggen, « The Interhexameric Contacts in the Four-hexameric Hemocyanin from the Tarantula Eurypelma californicum: A Tentative Mechanism for Cooperative Behavior », Journal of Molecular Biology, vol. 237, no 4, , p. 464-478 (PMID 8151706, DOI 10.1006/jmbi.1994.1248, lire en ligne)
- (en) Frank G. Perton, Jaap J. Beintema et Heinz Decker, « Influence of antibody binding on oxygen binding behavior of Panulirus interruptus hemocyanin », FEBS Letters, vol. 408, no 2, , p. 124-126 (PMID 9187351, DOI 10.1016/S0014-5793(97)00269-X, lire en ligne)
- (en) Heinz Decker et Felix Tuczek, « Tyrosinase/catecholoxidase activity of hemocyanins: structural basis and molecular mechanism », Trends in Biochemical Sciences, vol. 25, no 8, , p. 392-397 (PMID 10916160, DOI 10.1016/S0968-0004(00)01602-9, lire en ligne)
- (en) Heinz Decker, Thorsten Schweikardt, Dorothea Nillius, Uwe Salzbrunn, Elmar Jaenicke et Felix Tuczek, « Similar enzyme activation and catalysis in hemocyanins and tyrosinases », Gene, vol. 398, nos 1-2, , p. 183-191 (PMID 17566671, DOI 10.1016/j.gene.2007.02.051, lire en ligne)
- (en) David W. McFadden, Dale R. Riggs, Barbara J. Jackson et Linda Vona-Davis, « Keyhole limpet hemocyanin, a novel immune stimulant with promising anticancer activity in Barrett's esophageal adenocarcinoma », The American Journal of Surgery, vol. 186, no 5, , p. 552-555 (PMID 14599624, DOI 10.1016/j.amjsurg.2003.08.002, lire en ligne)