
Alternance de générations

L'alternance de générations (appelée aussi métagenèse) est une caractéristique du cycle de vie des plantes (prises ici au sens d'Archaeplastida). Une génération est un stade de développement multicellulaire issu d’un stade cellulaire (spore ou gamète) et produisant un stade cellulaire (gamète ou spore). La génération produisant des gamètes est le gamétophyte, et la génération produisant des spores est le sporophyte. De façon générale, un gamétophyte alterne avec un sporophyte. À maturité, le sporophyte produit des spores.
Les spores germent et se développent sous forme d'un gamétophyte. À maturité, le gamétophyte produit des gamètes par mitose, sans modification du nombre de chromosomes. Deux gamètes (provenant de deux organismes différents de la même espèce ou du même organisme) fusionnent pour produire un zygote, qui se développe sous la forme d'un sporophyte. Ce cycle, de gamétophyte à gamétophyte (ou bien de sporophyte à sporophyte), est la manière dont toutes les plantes terrestres et beaucoup d'algues subissent la reproduction sexuée.
Parce qu'elle constitue une étape-clé dans l'alternance des générations, il est probable que la méiose, processus qui permet de réduire de moitié le nombre de chromosomes, de 2n à n, ait une fonction adaptative fondamentale. La nature de cette fonction est encore en débat (voir Méiose), mais les deux idées principales sont que la méiose est adaptative parce qu'elle facilite la réparation des altérations qui peuvent affecter l'ADN ou parce qu'elle provoque des variations génétiques. Selon la place de la méiose dans le cycle, le sporophyte peut être haploïde ou diploïde, de même le gamétophyte peut être haploïde ou diploïde. Les places de la méiose et de la fécondation dans le cycle définissent les phases du cycle, et l’alternance des phases (haploïde et diploïde, voire dicaryotique lorsque la fusion des noyaux est déplacée dans le temps par rapport à la fusion des cytoplasmes lors de la fécondation). L’alternance cytologique des phases est indépendante de l’alternance morphologique des générations.
La relation entre le sporophyte et le gamétophyte varie selon les différents groupes de plantes. Chez les algues qui ont une alternance de générations, le sporophyte et le gamétophyte sont des organismes séparés indépendants, qui peuvent éventuellement avoir une apparence similaire. Chez les hépatiques, les mousses et les anthocérotes, le sporophyte est bien moins développé que le gamétophyte et est largement dépendant de ce dernier. Bien que les sporophytes des mousses et anthocérotes puissent assurer la photosynthèse, ils ont besoin de photosynthats supplémentaires provenant du gamétophyte pour soutenir leur croissance et le développement des spores, et en dépendent pour la fourniture d'eau, de nutriments minéraux et d'azote[1],[2]. Par contre, chez toutes les plantes vasculaires modernes, le gamétophyte est bien moins développé que le sporophyte, bien que leurs ancêtres du Dévonien avaient des gamétophytes et des sporophytes de complexité à peu près équivalente[3]. Chez les fougères, le gamétophyte est un petit prothalle autotrophe en forme de lame dont le jeune sporophyte est brièvement dépendant pour son alimentation. Chez les plantes à fleurs, la réduction du gamétophyte est extrême : il se compose de seulement quelques cellules qui se développent à l'intérieur du sporophyte.
Tous les animaux se développent différemment. Un animal adulte est diploïde et est donc, en un sens, l'équivalent d'un sporophyte. Toutefois, un animal produit « directement » des gamètes haploïdes par méiose. Il n'y a pas de spores haploïdes capables de se diviser, si bien qu'il y a pas d'équivalent d'un gamétophyte haploïde. Il n'y a pas d'alternance entre des formes diploïde et haploïde.
D'autres organismes, tels que les champignons, peuvent avoir des cycles de vie dans lesquels les différents types d'organismes alternent. Le terme d'« alternance de générations » s'applique également à ces cas.
Les cycles biologiques des plantes et des algues avec alternance de phases multicellulaires haploïdes et diploïdes sont dits « diplohaplontiques » (les termes équivalents « haplodiplontiques », ou « dibiontiques » sont également usités). Les cycles biologiques, tels que ceux des animaux, dans lesquels il n'y a qu'un seul stade multicellulaire diploïde sont appelés « diplontiques » (Les cycles biologiques dans lesquels il n'y a qu'un seul stade haploïde multicellulaire sont dits « haplontiques »).
Définition[modifier | modifier le code]
Dans l'exposé de « l'alternance de générations » ci-dessus, on considère que l'alternance d'une forme diploïde multicellulaire avec une forme haploïde multicellulaire est la caractéristique déterminante, indépendamment du fait que ces formes soient des organismes indépendants ou non[4]. Chez certaines espèces, telles que l'algue Ulva lactuca, les formes diploïdes et haploïdes sont en effet des organismes vivants indépendants, d'apparence pratiquement identique, que l'on qualifie donc d'isomorphes. Les gamètes haploïdes, nageant librement, forment un zygote diploïde qui germe pour produire un sporophyte diploïde multicellulaire. Le sporophyte produit par méiose des spores haploïdes, nageant librement, qui germent pour produire des gamétophytes haploïdes.
Cependant, chez certains autres groupes, le sporophyte ou le gamétophyte peuvent être fortement réduits et incapables de vivre librement. Par exemple, chez tous les bryophytes la génération du gamétophyte est dominante et le sporophyte est dépendant du gamétophyte. En revanche, chez toutes les plantes vasculaires terrestres modernes, les gamétophytes sont fortement réduits, bien que des témoins fossiles indiquent qu'elles dérivent d'ancêtres isomorphes[3]. Chez les plantes à graines, le gamétophyte femelle se développe totalement dans le sporophyte qui le protège et le nourrit ainsi que l'embryon du sporophyte qu'il produit. Les grains de pollen, qui sont les gamétophytes mâles, sont réduits à quelques cellules (trois cellules seulement dans de nombreux cas). Dans ce cas la notion de deux générations est moins évidente ; selon Bateman & Dimichele « [s]porophyte et gamétophyte fonctionnent en réalité comme un organisme unique »[5]. L'expression « alternance de phases » serait alors être plus appropriée[6].
Histoire[modifier | modifier le code]
Les débats sur l'alternance de générations au début du XXe siècle peuvent être source de confusion parce que plusieurs manières de classer les « générations » coexistaient (sexuée/asexuée, gamétophyte/sporophyte, haploïde/diploïde, etc.)[7].
Initialement, alors qu'ils étudiaient le développement des tuniciers, cnidaires et trématodes, Adelbert von Chamisso et Japetus Steenstrup ont décrit la succession des générations organisées différemment (sexuée et asexuée) chez les animaux sous le nom d'« alternance de générations »[7]. Ce phénomène est également connu sous le nom d'hétérogamie. Actuellement, l'expression « alternance de générations » est presque exclusivement associée aux cycles biologiques des plantes, en particulier avec l'alternance de gamétophytes et de sporophytes[7].
Wilhelm Hofmeister a démontré l'alternance morphologique de générations chez les plantes[8], entre une génération sporifère (sporophyte) et une génération porteuse de gamètes (gamétophyte)[9],[10]. À cette époque, a émergé un débat centré sur l'origine de la génération asexuée des plantes terrestres (c'est-à-dire le sporophyte), et classiquement caractérisé comme un conflit entre les théories de l'alternance de générations antithétique (Čelakovský, 1874) et homologue (Pringsheim, 1876)[7]. C'est Čelakovský qui a forgé les termes « sporophyte » et « gamétophyte ».
Eduard Strasburger (1874) a découvert l'alternance entre phases nucléaires diploïdes et haploïdes[7], appelée également « alternance cytologique de phases nucléaires »[11]. Bien qu'elles coïncident le plus souvent, l'alternance morphologique et l'alternance des phases nucléaires sont parfois indépendantes l'une de l'autre, par exemple, chez de nombreuses algues rouges, la même phase nucléaire peut correspondre à deux générations morphologiques différentes[11]. Chez certaines fougères qui ont perdu la reproduction sexuée, il n'y a aucun changement de phase nucléaire, mais l'alternance de générations est maintenue[12].
Alternance de générations chez les plantes[modifier | modifier le code]
Éléments fondamentaux[modifier | modifier le code]
Le diagramme ci-dessus montre les éléments fondamentaux de l'alternance de générations chez les plantes (il existe toutefois de nombreuses variantes chez les différents groupes de plantes). En partant de la droite du diagramme, les processus impliqués sont les suivants[13] :
- Deux gamètes haploïdes unicellulaires, contenant chacun n chromosomes non appariés, fusionnent pour former un zygote diploïde unicellulaire, qui contient dès lors n paires de chromosomes, soit 2n chromosomes au total.
- Le zygote diploïde unicellulaire germe, en se divisant par le processus normal (mitose), qui maintient le nombre de chromosomes à 2n. Le résultat est un organisme diploïde multi-cellulaire, appelée « sporophyte » (parce qu'à maturité il produit des spores).
- Le sporophyte mûr produit un ou plusieurs « sporanges » , organes qui produisent les cellules-mères des spores diploïdes (sporocystes). Celles-ci se divisent par un processus spécial (méiose) qui réduit le nombre de chromosomes de moitié. Cela se traduit d'abord dans quatre spores haploïdes unicellulaires, chacune contenant n chromosomes non appariés.
- La spore haploïde unicellulaire germe, en se divisant par le processus normal (mitose), ce qui maintient le nombre de chromosomes à n. Le résultat est un organisme haploïde multicellulaire, dit « gamétophyte » (parce que à maturité, il produit des gamètes).
- Le gamétophyte mûr produit un ou plusieurs « gamétanges », organes qui produisent les gamètes haploïdes. Au moins un type de gamètes possède un mécanisme lui permettant d'atteindre un autre gamète afin de fusionner avec lui.
L'« alternance de générations » dans le cycle biologique est donc l'alternance entre une génération de sporophytes diploïdes (2n) et une génération de gamétophytes haploïdes (n).

La situation est tout à fait différente de celle des animaux, chez lesquels le processus fondamental est qu'un individu diploïde (2n) produit « directement » des gamètes haploïdes (n) par la méiose. Les spores (c'est-à-dire des cellules haploïdes capables de subir la mitose) ne sont pas produites, de sorte qu'il n'existe aucun organisme multicellulaire haploïde. Les gamètes unicellulaires sont les seules entités haploïdes.
Hypothèses de mise en place de l'alternance des générations[modifier | modifier le code]
Deux hypothèses expliquent cette mise en place : la théorie de « l'intercalation » suppose que la méiose est retardée dans le cycle de développement. Une nouvelle génération diploïde s'intercale dans un cycle monogénétique haploïde ; la théorie de « la transformation » envisage que la génération sporophytique résulte de la transformation du gamétophyte qui est la génération originelle[14].
Variations[modifier | modifier le code]
Le diagramme présenté plus haut est une bonne représentation du cycle biologique de certaines algues multicellulaires (par exemple le genre Cladophora) qui ont des sporophytes et des gamétophytes d'apparence presque identique et qui n'ont pas des types différents de spores ou de gamètes[15].
Cependant, il existe beaucoup de variations possibles des éléments fondamentaux d'un cycle biologique avec alternance des générations. Chaque variation peut se produire séparément ou en combinaison, ce qui conduit à une diversité déconcertante de cycles biologiques. Les termes utilisés par les botanistes pour décrire ces cycles biologiques peuvent aussi être déroutants. Comme l'ont dit Bateman et Dimichele « […] l'alternance de générations est devenue un bourbier terminologique ; souvent, un terme représente plusieurs concepts ou un concept est représenté par plusieurs termes »[16].
Les variations possibles sont les suivantes :
- Importance relative du sporophyte et du gamétophyte.
- Égale (homomorphie ou isomorphie).
Les algues filamenteuses du genre Cladophora, qui vivent principalement en eau douce, ont des sporophytes diploïdes et des gamétophytes haploïdes qui sont impossibles à distinguer par leur aspect extérieur[17].
Aucune plante terrestre vivant actuellement n'a de sporophytes ou gamétophytes également dominants, bien que certaines théories de l'évolution de l'alternance des générations suggèrent que c'étaient le cas chez leurs ancêtres. - Inégale (hétéromorphie ou anisomorphie).

Gamétophyte d'une mousse, Mnium hornum. - Gamétophyte dominant (gamétophytique).
Chez les hépatiques, mousses et anthocérotes, la forme dominante est le gamétophyte haploïde. Le sporophyte diploïde n'a pas d'existence indépendante, tirent l'essentiel des éléments nutritifs du gamétophyte parent, et n'ayant pas de chlorophylle à maturité[18].
Sporophyte d'une fougère, Blechnum discolor. - Sporophyte dominant (sporophytique).
Chez les fougères, tant le sporophyte que le gamétophyte sont capables de vivre de façon indépendante, mais la forme dominante est le sporophyte diploïde. Le gamétophyte haploïde est beaucoup plus petit et de structure plus simple. Chez les plantes à graines, le gamétophyte est encore plus réduit (avec au minimum trois cellules seulement), tirant tous ses éléments nutritifs du sporophyte.
La réduction extrême de la taille du gamétophyte et sa rétention dans le sporophyte signifie que lorsqu'on l'applique aux plantes à graines, l'expression « alternance de générations » est quelque peu fallacieuse : « [s]porophyte et gamétophyte fonctionnent en réalité comme un seul organisme »[5]. Certains auteurs préfèrent l'expression « alternance de phases »[6].
- Gamétophyte dominant (gamétophytique).
- Égale (homomorphie ou isomorphie).
- Différenciation des gamètes.
- Gamètes identiques (isogamie).
Comme d'autres espèces du genre Cladophora, C. callicoma a des gamètes flagellés qui sont d'aspect identique et en capacité de mouvement[17]. - Gamètes de deux tailles différentes (anisogamie).
- Les deux ont une motilité similaire.
Les espèces d'Ulva (laitue de mer) ont des gamètes qui ont tous deux flagellés et sont donc mobiles. Cependant, ils sont de deux tailles différentes : des gamètes « femelles » plus grands et des gamètes « mâles » plus petits[19]. - Un gamète grand et fixe, l'autre petit et mobile (oogamie).
Les mégagamètes fixes, plus grands, sont les gamètes femelles (ovules), et les microgamètes mobiles, plus petits, sont les gamètes mâles (spermatozoïdes). Le degré de la motilité des spermatozoïdes peut être très limité (comme dans le cas de plantes à fleurs) mais tous sont en mesure de se déplacer vers les ovules fixes. Lorsque (ce qui est presque toujours le cas), les spermatozoïdes et les ovules sont produits dans deux types de gamétanges différents, ceux qui produisent des spermatozoïdes sont appelées « anthéridies » et ceux qui produisent des ovules, « archégones ».
Gamétophyte de Pellia epiphylla avec sporophytes croissant à partir des vestiges d'archégones. - Anthéridies et archégones formés sur le même gamétophyte, qui est alors qualifié de « monoïque ».
Chez l'hépatique, Pellia epiphylla, le gamétophyte est la génération dominante. Elle est monoïque : les petites anthéridies productrices de spermatozoïdes rougeâtres sont dispersées le long de la nervure centrale, tandis que les archégones productrices d'ovules poussent plus près des extrémités des divisions de la plante[20]. - Anthéridies et archégones formés sur des gamétophytes différents, qui sont appelés « dioïques ».
Chez la mousse, Mnium hornum, le gamétophyte est la génération dominante. Elle est « dioïque » : les plants mâles produisent seulement des anthéridies en rosettes terminales, les plants femelles seulement des archégones en forme de capsules pédonculées[21]. Les gamétophytes des plantes à graines sont aussi « dioïque ». Cependant le sporophyte parent peut être « monoïque », produisant à la fois des gamétophytes mâles et femelles, ou « dioïque », produisant des gamétophytes d'un seul genre.
Les gamétophytes des plantes à graines sont de taille extrêmement réduite ; l'archégone se compose d'un petit nombre de cellules, et le gamétophyte mâle peut être constitué de deux cellules seulement[22].
- Anthéridies et archégones formés sur le même gamétophyte, qui est alors qualifié de « monoïque ».
- Les deux ont une motilité similaire.
- Gamètes identiques (isogamie).
- Différenciation des spores.
- Toutes les spores de même taille (homosporie ou isosporie).
Les prêles (genre Equisetum) ont des spores qui sont toutes de la même taille[23]. - Spores de deux tailles distinctes (hétérosporie ou anisosporie) : mégaspores et microspores.
Lorsque les deux types de spores sont produites dans des sporanges de types différents, ceux-ci sont appelés « mégasporanges » et « microsporanges ». Une mégaspore se développe souvent (mais pas toujours) aux dépens des trois autres cellules issues de la méiose, qui avortent.- Mégasporanges et microsporanges formés sur le même sporophyte, qui est alors appelé « monoïque ».
La plupart des plantes à fleurs appartiennent à cette catégorie. Ainsi la fleur du lis contient six étamines (microsporanges) produisant des microspores qui se développent en grains de pollen (microgamétophytes), et trois carpelles fusionnés qui produisent des mégasporanges sans tégument (ovules) dont chacun produit un mégaspore qui se développe dans le mégasporange pour former le mégagamétophyte.
Chez d'autres plantes, telles que le noisetier, certaines fleurs n'ont que des étamines, d'autres que des carpelles, mais la même plante (c'est-à-dire le sporophyte) porte les deux types de fleurs et est donc monoïque.
Fleurs de houx (espèce dioïque) : mâles en haut, femelles en bas (feuilles coupées pour mieux montrer les fleurs). - Mégasporanges et microsporanges formés sur des sporophytes différents, qui sont alors appelés « dioïques »
Un pied de houx (Ilex aquifolium) porte soit des fleurs « mâles » qui possèdent seulement des étamines fonctionnelles (microsporanges) produisant des microspores qui se développent en grains de pollen (microgamétophytes) ou bien des fleurs « femelles » qui ne possèdent que des carpelles fonctionnels produisant des mégasporanges (ovules) sans tégument contenant une mégaspore qui se développe en un mégagamétophyte multicellulaire.
- Mégasporanges et microsporanges formés sur le même sporophyte, qui est alors appelé « monoïque ».
- Toutes les spores de même taille (homosporie ou isosporie).




Il existe des corrélations entre ces variations, mais ce ne sont que des corrélations, et pas des liens absolus. Par exemple, chez les plantes à fleurs, les microspores produisent finalement des microgamètes (mâles) et les mégaspores produisent finalement des mégagamètes (femelles). Cependant, chez les fougères et leurs alliés, il existe des groupes ayant des spores indifférenciées mais avec des gamétophytes différenciées. Par exemple, la fougère, Ceratopteris thalictrioides, a des spores d'un seul type, qui varie continûment en taille. Les spores les plus petites tendent à germer en gamétophytes ne produisant que des anthéridies productrices de spermatozoïdes[23].
Un cycle biologique complexe[modifier | modifier le code]

Le diagramme ci-contre montre l'alternance de générations chez une espèce qui est hétéromorphe, sporophytique, oogamique, hétérosporique et dioïque. Un exemple de plante à graines pourrait être le saule (la plupart des espèces du genre Salix sont dioïques)[24].
En partant du centre du diagramme, les processus impliqués sont les suivants :
- Une oosphère immobile, contenue dans un archégone, fusionne avec un spermatozoïde mobile, libéré par une anthéridie. Le zygote résultant est soit mâle soit femelle.
- Un zygote mâle se développe par mitose en un microsporophyte, qui produit à maturité un ou plusieurs microsporanges. Les microspores se développent à l'intérieur du microsporange par méiose
Chez le saule (comme chez toutes les plantes à graines) le zygote se développe d'abord dans un microsporophyte embryonnaire au sein de l'ovule (mégasporange enfermé dans une ou plusieurs couches de tissu protectrice, le tégument). À maturité, ces structures deviennent la graine. Plus tard, la graine tombe, germe et se développe pour former un arbre adulte. Un saule « mâle » (microsporophyte) produit des fleurs portant seulement des étamines, dont les anthères sont les microsporanges. - Les microspores germent en produisant des microgamétophytes ; à maturité une ou plusieurs anthéridies sont formées. Les anthérozoïdes ou spermatozoïdes se développent dans l'anthéridie.
Chez le saule, les microspores ne sont pas libérées par l'anthère (le microsporange), mais se développent dans les grains de pollen (microgamétophytes). Tout le grain de pollen est transporté (par un insecte ou par le vent) vers un ovule (mégagamétophyte), où un spermatozoïde est produit et descend dans le tube pollinique pour atteindre l'oosphère. - Un zygote « femelle » se développe par mitose en un mégasporophyte, qui produit à maturité un ou plusieurs mégasporanges. Les mégaspores se développent dans les mégasporanges ; généralement l'une des quatre spores produites par méiose grossit au détriment des trois autres, qui disparaissent.
Les saules « femelles » (mégasporophytes) produisent des fleurs ayant seulement des carpelles (feuilles modifiées portant les mégasporanges). - Les mégaspores germent en produisant des mégagamétophytes ; à maturité, un ou plusieurs archégones sont produits. Les oosphères se développent à l'intérieur des archégones.
Les carpelles d'un saule produisent des ovules, les mégasporanges protégés par des téguments. Dans chaque ovule, une mégaspore se développe par mitose en un mégagamétophyte. Un archégone se développe dans le mégagamétophyte et produit une oosphère. L'ensemble de la « génération » gamétophytique reste sous la protection du sporophyte, à l'exception des grains de pollen (qui ont été réduits à seulement trois cellules contenues dans la paroi du microspore).
- Un zygote mâle se développe par mitose en un microsporophyte, qui produit à maturité un ou plusieurs microsporanges. Les microspores se développent à l'intérieur du microsporange par méiose
Cycles biologiques des différents groupes de plantes[modifier | modifier le code]
Le terme « plante » est pris ici au sens d'Archaeplastida, c'est-à-dire les glaucophytes, les algues rouges et vertes et les plantes terrestres.
L'alternance de générations se produit chez presque toutes les algues rouge et vertes multicellulaires, tant chez les formes d'eau douce (comme Cladophora ) que chez les algues marines (comme Ulva). En général, les générations sont homomorphes (isomorphes) et ont une existence autonome. Certaines espèces d'algues rouges ont une alternance de générations complexe, à trois phases, dont une phase de gamétophyte et deux phases distinctes de sporophyte.
Les plantes terrestres ont toutes une alternance de générations hétéromorphe (anisomorphe), dans lequel le sporophyte et le gamétophyte sont nettement différents.
Chez les embryophytes, le cycle devient de plus en plus hétéromorphe avec « une réduction de plus en plus importante du gamétophyte (aussi bien en taille qu'en durée de vie) : Bryophytes, Ptéridophytes de type Polypodium, Selaginella, Marsilea, Spermatophytes où il devient microscopique). Cela va de pair avec la sexualisation du sporophyte. D'abord nulle (un seul type de spores chez Funaria (en) et Polypodium), puis faible (tendance à l'existence de deux types de spores chez Equisetum), moyenne (deux types de sporanges et de spores chez les Ptéridophytes hétérosporés), forte chez les plantes à fleurs hermaphrodites et monoïques (étamines et carpelles) et enfin totale chez les plantes dioïques où la séparation des sexes est permanente. Cette réduction progressive s'accompagne d'une protection de plus en plus grande de la phase haploïde[25] ».
Chez tous les bryophytes (hépatiques, mousses et anthocérotes) la génération des gamétophytes est la plus visible.
À titre d'illustration, considérons une mousse monoïque. Les anthéridies et archégones se développent sur la plante mûre (le gamétophyte). En présence d'eau, le spermatozoïde biflagellé issu des anthéridies rejoint l'archégone en nageant et la fécondation a lieu, permettant le développement d'un sporophyte diploïde. Le sporophyte croît à partir de l'archégone. Sa structure se compose d'une longue tige surmontée d'une capsule à l'intérieur de laquelle les cellules produisant les spores subissent la méiose pour former des spores haploïdes. La plupart des mousses comptent sur le vent pour disperser ces spores, bien qu'une espèce comme Splachnum sphaericum (en) soit entomophile, faisant appel à des insectes pour disperser ses spores.
-

Alternance de générations chez les hépatiques. -

Cycle biologique des mousses. -

Cycle biologique des anthocérotes.



Chez les fougères et de leurs alliés, y compris les lycopodes et les prêles, l'organisme le plus remarquable observé sur le terrain est le sporophyte diploïde. Les spores haploïdes se développent dans des sores à la face inférieure des frondes et sont dispersées par le vent (parfois en flottant sur l'eau). Si les conditions sont favorables, une spore va germer et former un organisme plutôt discret appelé prothalle. Le prothalle haploïde ne ressemble pas au sporophyte, et de ce fait les fougères et leurs alliés ont une alternance de générations hétéromorphe. Le prothalle a une courte durée de vie, mais c'est lui qui réalise la reproduction sexuée, conduisant à la formation du zygote diploïde, qui pousse alors à partir du prothalle en tant que sporophyte.
-
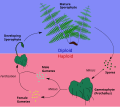
Alternance de générations chez les fougères. -

Gamétophyte (prothalle) de Dicksonia sp. -

Sporophyte de Dicksonia antarctica. -

Sores à la face inférieure d'une fronde de Dicksonia antarctica.




Chez les spermatophytes, ou plantes à graines, le sporophyte est la phase multicellulaire dominante ; les gamétophytes sont fortement réduits en taille et de morphologie très différente. La totalité de la génération du gamétophyte, à la seule exception des grains de pollen (microgamétophytes), est contenue dans le sporophyte. Le cycle biologique d'une plante à fleurs (angiosperme) dioïque , le saule, a été décrit plus haut (Un cycle biologique complexe). Le cycle biologique d'un gymnosperme est similaire. Cependant, les plantes à fleurs ont en plus un phénomène appelé « double fécondation ». Deux noyaux de spermatozoïdes provenant d'un grain de pollen (microgamétophyte), au lieu d'un seul spermatozoïde, pénètrent dans l'archégone du mégagamétophyte ; l'un fusionne avec le noyau de l'oosphère pour former le zygote, les autres fusionnent avec deux autres noyaux du gamétophyte pour former l'endosperme, qui nourrit l'embryon au cours de son développement.
Émergence évolutive de la phase diploïde dominante[modifier | modifier le code]

Une hypothèse est que la base de l'émergence de la phase diploïde du cycle biologique (sporophyte) comme phase dominante (par exemple chez les plantes vasculaires) pourrait être que la diploïdie autorise le masquage de l'expression de mutations délétères par complémentation génétique[26],[27]. Ainsi, si l'un des génomes parentaux dans les cellules diploïdes contenait des mutations conduisant à des défauts dans un ou plusieurs produits géniques, ces lacunes pourraient être compensées par l'autre génome parental (qui pourrait néanmoins avoir ses propres défauts dans d'autres gènes). À mesure que la phase diploïde devenait prédominante, l'effet de masquage a sans doute permis l'accroissement de la taille du génome, et donc du contenu en information, sans la contrainte d'avoir une meilleure précision de la réplication de l'ADN. La possibilité d'augmenter le contenu en information à faible coût fut avantageuse, car elle a permis d'encoder de nouvelles adaptations. Ce point de vue a été contesté, avec des preuves montrant que la sélection est plus efficace dans les phases haploïdes que dans les phases diploïdes du cycle biologiques des mousses et des angiospermes[28].
Toujours est-il que le cycle de développement diphasique avec dominance du sporophyte a été sélectionné au cours de l'évoolution. De plus, chez les Spermatophytes, le gamétophyte est miniaturisé : le gamétophyte mâle (contenu du grain de pollen) et femelle (sac embryonnaire) sont devenus microscopiques.
-

Cycle biologique des Angiospermes. -

Pollen (microgamétophytes) à l'extrémité d'une étamine de tulipe. -

Ovules (mégagamétophytes) : Gymnospermes à gauche, Angiospermes à droite. -

Double fécondation.




Processus similaires chez d'autres organismes[modifier | modifier le code]
Chromalveolata[modifier | modifier le code]
Certains organismes classés dans les Chromalveolata, qui ne sont donc pas des plantes au sens utilisé dans cet article, présentent une alternance de générations. Les Foraminifères subissent une alternance de générations hétéromorphe entre les formes haploïde, « Gamontes », et diploïde, « Schizontes ». L'organisme haploïde unicellulaire est généralement beaucoup plus grand que l'organisme diploïde.
Champignons (Fungi)[modifier | modifier le code]
Les mycéliums fongiques sont généralement haploïdes.
Lorsque des mycéliums de types sexuels différents se rencontrent, ils produisent deux cellules multinucléées en forme de balle, qui se rejoignent via un « pont d'accouplement ». Les noyaux se déplacent d'un mycélium à l'autre, formant un « hétérocaryon » (« différents noyaux »). Ce processus est appelé « plasmogamie ».
La fusion réelle pour former des noyaux diploïdes est appelée «caryogamie », et ne peut pas se produire avant que les sporanges soient formés. La caryogamie produit un zygote diploïde, sporophyte éphémère qui subit rapidement la méiose pour former des spores haploïdes. Lorsque les spores germent, elles se développent en nouveaux mycéliums.
Myxomycètes[modifier | modifier le code]
Le cycle biologique des myxomycètes est très similaire à celui des champignons. Les spores haploïdes germinent pour former des cellules d'essaimage ou « myxamoebae ». Ils fusionnent dans un processus appelé « plasmogamie » et « caryogamie » pour former un zygote diploïde. Le zygote se développe en un plasmode qui, arrivé à maturité, produit selon les espèces une ou plusieurs fructifications contenant des spores haploïdes.
Animaux[modifier | modifier le code]
L'alternance entre générations diploïde multicellulaire et haploïde multicellulaire ne se rencontre jamais chez les animaux[29]. Chez certains animaux, il existe une alternance entre phases de parthénogénèse et de reproduction sexuelle (hétérogamie). Les deux phases sont diploïdes. Ce processus a parfois été appelé « alternance de générations »[30], mais c'est tout à fait différent.
Notes et références[modifier | modifier le code]
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Alternation of generations » (voir la liste des auteurs).
- (en) R.J. Thomas, D.S. Stanton, D.H. Longendorfer et M.E. Farr, « Physiological evaluation of the nutritional autonomy of a hornwort sporophyte », Botanical Gazette, vol. 139, no 3, , p. 306–311 (DOI 10.1086/337006).
- (en) J.M. Glime, « Ecophysiology of Development: Sporophyte. Chap t. 5-9 », dans Bryophyte Ecology: Vol. 1 Physiological Ecology, Michigan Technological University and the International Association of Bryologists, (lire en ligne).
- (en) H. Kerp, N.H. Trewin et H. Hass, « New gametophytes from the Lower Devonian Rhynie Chert », Transactions of the Royal Society of Edinburgh: Earth Sciences, vol. 94, no 4, , p. 411–428 (DOI 10.1017/S026359330000078X).
- Taylor, Kerp et Hass 2005.
- Bateman et Dimichele 1994, p. 403.
- .Stewart et Rothwell 1993.
- (en) David Haig, « Homologous versus antithetic alternation of generations and the origin of sporophytes », The Botanical Review, vol. 74, no 3, , p. 395–418 (DOI 10.1007/s12229-008-9012-x, lire en ligne).
- (en) Nils Svedelius, « Alternation of Generations in Relation to Reduction Division », Botanical Gazette, vol. 83, no 4, , p. 362–384 (DOI 10.2307/2470766, JSTOR 2470766).
- (de) W. Hofmeister, Vergleichende Untersuchungen der Keimung, Entfaltung und Fruchtbildildiung höherer Kryptogamen (Moose, Farne, Equisetaceen, Rhizocarpeen und Lycopodiaceen) und der Samenbildung der Coniferen, Leipzig, F. Hofmeister, (lire en ligne). Traduction en anglais : (en) Frederick Currey, On the germination, development, and fructification of the higher Cryptogamia, and on the fructification of the Coniferæ, Londres, Robert Hardwicke, (lire en ligne)
- (fr) J. Feldmann et G. Feldmann, « Recherches sur les Bonnemaisoniacées et leur alternance de générations », Annales des Sciences Naturelles, Botanique, série 11, vol. 3, , p. 75–175 (lire en ligne), p. 157.
- (fr) J. Feldmann, « Les problèmes actuels de l'alternance de générations chez les Algues », Bulletin de la Société Botanique de France, vol. 119, , p. 7–38 (DOI 10.1080/00378941.1972.10839073, lire en ligne).
- (en) P. Schopfer, H. Mohr, « Physiology of Development », dans Plant physiology, Berlin, Springer, , 288–291 p. (ISBN 3-540-58016-6, lire en ligne).
- Sauf indication contraire, les informations contenues dans cette section proviennent de Foster et Gifford 1974, Sporne 1974a et Sporne 1974b.
- Daniel Richard, Patrick Chevalet, Nathalie Giraud, Fabienne Pradere, Thierry Soubaya, Sciences de la vie pour le Capes et l'Agrégation, Dunod, (Modèle:Google Livvres), p. 126.
- Guiry et Guiry 2008.
- Bateman et Dimichele 1994, p. 347.
- Shyam 1980.
- Watson 1981, p. 2.
- Kirby 2001.
- Watson 1981, p. 425–6.
- Watson 1981, p. 287–8.
- Sporne 1974a, p. 17–21.
- Bateman et Dimichele 1994, p. 350–1.
- (en) Encyclopædia Britannica, vol. XIX, New York, Encyclopædia Britannica, , 11e éd. (lire en ligne).
- Yves Tourte, Michel Bordonneau, Max Henry et Catherine Tourte, Le monde des végétaux. Organisation, physiologie et génomique, Dunod, (lire en ligne), p. 48
- (en) H. Bernstein, G.S. Byers et R.E. Michod, « Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation », The American Naturalist, vol. 117, no 4, , p. 537–549 (DOI 10.1086/283734).
- (en) R.E. Michod et T.W. Gayley, « Masking of mutations and the evolution of sex », The American Naturalist, vol. 139, no 4, , p. 706–734 (DOI 10.1086/285354).
- (en) Péter Szövényi, Mariana Ricca, Zsófia Hock, Jonathan A. Shaw, Kentaro K. Shimizu et Andreas Wagner, « Selection is no more efficient in haploid than in diploid life stages of an angiosperm and a moss », Molecular Biology and Evolution, vol. 30, no 8, , p. 1929 (PMID 23686659, DOI 10.1093/molbev/mst095).
- Barnes et al. 2001, p. 321.
- Scott 1996, p. 35.
Voir aussi[modifier | modifier le code]
Articles connexes[modifier | modifier le code]
- Cycle de vie (biologie)
- Histoire évolutive des végétaux#Évolution des cycles de vie : origine évolutive de l'alternance de phases.
Bibliographie[modifier | modifier le code]
- (en) R.S.K. Barnes, P. Calow, P.J.W. Olive, D.W. Golding et J.I. Spicer, The Invertebrates : a synthesis, Oxford; Malden, MA, Blackwell, , 512 p. (ISBN 978-0-632-04761-1)
- (en) R.M. Bateman et W.A. Dimichele, « Heterospory – the most iterative key innovation in the evolutionary history of the plant kingdom », Biological Reviews of the Cambridge Philosophical Society, vol. 69, , p. 315–417 (lire en ligne)
- (en) P.R. Bell et A.R. Hemsley, Green Plants : their Origin and Diversity, Cambridge, etc., Cambridge University Press, , 2e éd., 360 p. (ISBN 978-0-521-64109-8)
- (en) A.S. Foster et E.M. Gifford, Comparative Morphology of Vascular Plants, San Francisco, W.H. Freeman, , 2e éd. (ISBN 978-0-7167-0712-7)
- (en) M.D. Guiry et G.M. Guiry, « Cladophora », sur AlgaeBase, World-wide electronic publication, National University of Ireland, Galway,
- (en) A. Kirby, « Ulva, the sea lettuce », Monterey Bay Aquarium Research Institute,
- (en) Thomas Scott, Concise Encyclopedia Biology, Berlin, Walter de Gruyter, , 1287 p. (ISBN 978-3-11-010661-9)
- (en) R. Shyam, « On the life-cycle, cytology and taxonomy of Cladophora callicoma from India », American Journal of Botany, vol. 67, no 5, , p. 619–24 (DOI 10.2307/2442655, JSTOR 2442655)
- (en) K.R. Sporne, The Morphology of Angiosperms, Londres, Hutchinson, 1974a (ISBN 978-0-09-120611-6)
- (en) K.R. Sporne, The Morphology of Gymnosperms, Londres, Hutchinson, 1974b, 2e éd., 216 p. (ISBN 978-0-09-077152-3)
- (en) W.N. Stewart et G.W. Rothwell, Paleobotany and the Evolution of Plants, Cambridge, Royaume-Uni, Cambridge University Press, , 2e éd., 521 p. (ISBN 978-0-521-38294-6, lire en ligne)
- (en) E.V. Watson, British Mosses and Liverworts : An Introductory Work, Cambridge, Royaume-Uni, Cambridge University Press, , 3e éd., 519 p. (ISBN 978-0-521-28536-0, lire en ligne)
- (en) T.N. Taylor, H. Kerp et H. Hass, « Life history biology of early land plants: Deciphering the gametophyte phase », Proceedings of the National Academy of Sciences of the United States of America, vol. 102, no 16, , p. 5892–5897 (PMID 15809414, PMCID 556298, DOI 10.1073/pnas.0501985102).