
Membre chiridien
Cet article est une ébauche concernant l’anatomie et la biologie.
Vous pouvez partager vos connaissances en l’améliorant (comment ?) selon les recommandations des projets correspondants.
Consultez la liste des tâches à accomplir en page de discussion.


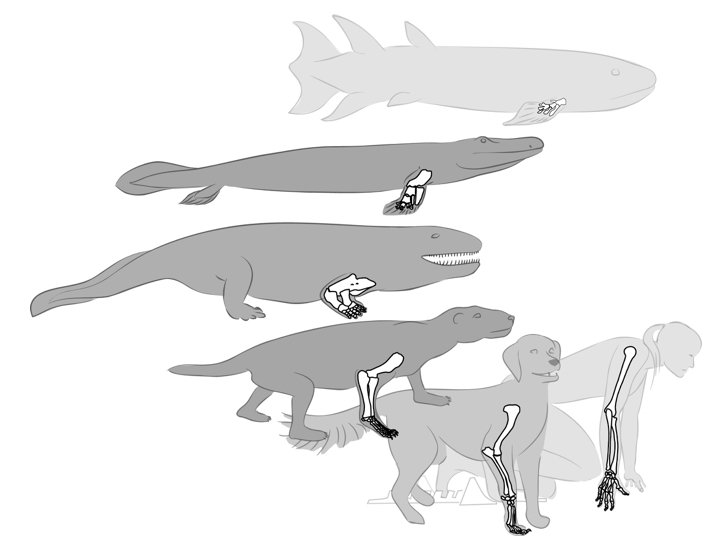
Les membres chiridiens (du grec ancien χείρ (kheir) : « main ») sont les appendices locomoteurs articulés et munis de doigts des tétrapodes, dérivant probablement de deux paires des nageoires de leurs ancêtres sarcoptérygiens ichtyens. Cette innovation anatomique apparue au Dévonien (passage de la nageoire des sarcoptérygyens tétrapodomorphes au membre chiridien des tétrapodes)[5] sert aux premiers tétrapodes aquatiques à nager et marcher sur le fond d'eaux saumâtres (marais côtiers, estuaires, lagunes et autres milieux aquatiques de salinité variable) avant de permettre la mise en place de la quadrupédie, mode de locomotion terrestre initial largement privilégié au sein des vertébrés terrestres[6],[7].
Ils sont originellement au nombre de deux paires articulées comme les nageoires sur une ceinture pectorale et pelvienne, mais peuvent être perdus secondairement, soit en partie (Amphiuma, Chalcides striatus), soit totalement (apodie chez les serpents, les gymnophiones…). Cependant, contrairement au membre ptérygien hétérogène chez les poissons, le membre chiridien (appelé aussi chiridium ou cheiropterygium) constitue un ensemble homogène[8].
Les tétrapodes possèdent deux paires de membres chiridiens dont les os sont mis en mouvement par la force exercée par les tendons à la suite de la contraction des muscles. Ce groupe rassemble aujourd'hui les lissamphibiens (seul groupe survivant de la classe des amphibiens) aux doigts nus et des amniotes (reptiles, oiseaux, mammifères) qui possèdent des étuis cornés couvrant les phalanges terminales des doigts, les griffes (formant notamment des sabots chez les ongulés, et des ongles chez les primates)[9].
Structure
[modifier | modifier le code]
Le membre chiridien est caractérisé par 3 segments articulés[10] :
- le segment proximal ou stylopode qui correspond, pour le membre supérieur, au bras (stricto sensu) comportant l’humérus, et pour le membre inférieur, à la cuisse comportant le fémur ;
- le segment moyen ou zeugopode (ou encore zygopode) qui correspond à l’avant-bras, comportant l’ulna et le radius, ou à la jambe (stricto sensu), comportant le tibia et la fibula. Ces os constitutifs du zeugopode peuvent être secondairement soudés (ex : les équidés, la jambe du hérisson). Chez certains afrothériens, notamment la taupe dorée, le zeugopode comporte non pas deux os mais trois ;
- le segment distal ou autopode qui correspond à la main ou au pied. Complexe, il se subdivise en trois sous-régions anatomiques :
- le basipode (proximal) qui comprend les carpes (poignet chez l'humain, genou chez les équidés) ou les tarses (cheville chez l'humain, jarret chez les équidés),
- le métapode (intermédiaire) qui correspond au métacarpe ou au métatarse,
- l’acropode qui correspond aux phalanges des doigts ancestraux (polydactylie puis pentadactylie).
Évolution
[modifier | modifier le code]



Chez les premiers tétrapodes terrestres, les membres chiridiens constituaient initialement les pattes, permettant la marche quadrupède. Au sein des différentes familles de Tétrapodes, le plan d'organisation général des membres à trois segments est hautement conservé (les membres antérieurs et postérieurs sont ainsi homologues aux bras et jambes humains), mais on observe des variations (modifications dans les proportions des différents os, fusions, réductions ou disparitions de structures) selon les espèces. À la suite de changements radicaux de niche écologique, ces membres ont pu se spécialiser pour permettre différents modes de locomotion. La concordance des morphologies obtenues constitue un exemple typique de convergence évolutive[7].
L'adaptation à la course quadrupède est réalisée essentiellement chez les Mammifères, à membre dressé parasagittal[16], par trois modifications concomitantes : l'allongement des membres qui intéresse surtout le métapode mais aussi celle des os proximaux relativement à ceux du segment moyen[17], le relèvement progressif de l'autopode (évolution dans le sens plantigrade → digitigrade → onguligrade), et la réduction progressive du nombre des doigts (perte des phalanges latérales par rapport à l'extrémité pentadactyle ancestrale)[18].
Les membres chiridiens (surtout antérieurs) se sont transformés en ailes pour permettre le vol chez les Ptérosaures (aujourd’hui disparus), les Oiseaux et les Chiroptères. La structure adoptée diffère pour chacun de ces groupes par le nombre de doigts supportant l'aile (respectivement un, deux et quatre).
Au cours de l'évolution plusieurs groupes de tétrapodes ont adopté un mode de vie aquatique ou semi-aquatique de telle sorte que leurs membres ont évolué en palettes natatoires :
- sauropsides fossiles
- Mésosauridés†, Sauroptérygiens†, Mosasaures†, Ichtyosaures†, Thalattosauriens† ...
- mammifères
- Cétacés, Siréniens, Pinnipèdes, Lutrinae, Castoridae ...
- oiseaux
- Sphenisciformes, Ansériformes, Pelecanidae ...
- anapsides
- Chéloniidés
Le membre antérieur chiridien chez les bipèdes[19] ne présente pas de caractères anatomiques évolutifs bien spécifiques[réf. souhaitée]. La plupart des théropodes du Mésozoïque avaient des membres antérieurs extrêmement courts qu'on suspecte non-fonctionnels. Chez les oiseaux, seuls théropodes du Cénozoïque, les membres antérieurs se sont adaptés en ailes, mais ont pu subséquemment chez certains groupes, après la perte de l'aptitude au vol, s'atrophier. Cette atrophie a abouti à une disparition complète chez les Dinornithiformes†. Les autres oiseaux non-volants, les ratites, ont conservé leurs ailes au moins sous forme vestigiale.
La bipédie existe aussi chez certains primates, essentiellement les descendants d'Homo Erectus†, et en particulier chez Homo Sapiens, pour qui les membres antérieurs ont perdu toute fonction locomotrice et se sont spécialisés pour la préhension et, selon certaines analyses[20], pour le lancer.
Reptation
[modifier | modifier le code]l'apodie est complète chez les serpents[21] et chez les gymnophiones du fait de la reptation.
-
 Homologie des membres antérieurs de vertébrés.
Homologie des membres antérieurs de vertébrés. -
![Relèvement progressif de l'autopode dans l'adaptation à la course quadrupède (membre antérieur : de gauche à droite : homme plantigrade, chien digitigrade et cheval onguligrade)[22].](//upload.wikimedia.org/wikipedia/commons/thumb/1/18/Les_diff%C3%A9rents_types_de_locomotions_chez_les_amniotes.png/120px-Les_diff%C3%A9rents_types_de_locomotions_chez_les_amniotes.png) Relèvement progressif de l'autopode dans l'adaptation à la course quadrupède (membre antérieur : de gauche à droite : homme plantigrade, chien digitigrade et cheval onguligrade)[22].
Relèvement progressif de l'autopode dans l'adaptation à la course quadrupède (membre antérieur : de gauche à droite : homme plantigrade, chien digitigrade et cheval onguligrade)[22].

![Relèvement progressif de l'autopode dans l'adaptation à la course quadrupède (membre antérieur : de gauche à droite : homme plantigrade, chien digitigrade et cheval onguligrade)[22].](/wiki/Fichier:Les_diff%C3%A9rents_types_de_locomotions_chez_les_amniotes.png)
Notes et références
[modifier | modifier le code]- A. Eusthenopteron ; B. Gogonasus ; C. Panderichthys.
- Le lobe musculaire de la nageoire est frangé de rayons dermiques, pièces exosquelettiques osseuses allongées qui soutiennent la partie périphérique membranaire, non musculaire de la nageoire.
- Les membres antérieurs d'Acanthostega (E) ont huit doigts, ceux d'Ichthyostega (F) et Tulerpeton (G) en ont six
- André Beaumont, Pierre Cassier, Daniel Richard, Biologie animale. Les Cordés : Anatomie comparée des vertébrés, Dunod, (lire en ligne), p. 203-204.
- (en) Mikiko Tanaka, « Fins into limbs: Autopod acquisition and anterior elements reduction by modifying gene networks involving 5’Hox, Gli3, and Shh », Developmental Biology, vol. 413, no 1, , p. 1-7 (DOI 10.1016/j.ydbio.2016.03.007).
- (en) Catherine A. Boisvert, « The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion », Nature, vol. 438, no 7071, , p. 1145–1147 (DOI 10.1038/nature04119).
- Lewis Wolpert, Cheryll Tickle, Alfonso Martinez Arias, Biologie du développement. Les grands principes, Dunod, , p. 639-641.
- Membres. Les Grands Articles d'Universalis, Encyclopaedia Universalis, (lire en ligne), n.p.
- André Beaumont, Pierre Cassier, Daniel Richard, Biologie animale. Les Cordés, Dunod, , p. 123.
- Christiane Perrier, Jean-François Beaux (dir.), Biologie-Géologie, Dunod, , p. 173
- Le pouce libre est utile « durant le vol pour positionner le bord d'attaque de la voilure, tout comme pour les déplacements à pied. L'ancrage du pouce est facilité par sa griffe ». Cf Laurent Arthur, Michèle Lemaire, Les Chauves-souris de France Belgique Luxembourg et Suisse, éditions Biotope, (lire en ligne), n.p.
- Membres ancestraux qui se plient en Z et sont situés de chaque côté du tronc, caractéristiques de la posture tentaculaire chez les Amphibiens du Primaire, les Urodèles et Reptiles actuels.
- Membres de type dressé, situés sous le tronc, caractéristiques de la posture érigée des Mammifères et des Tétrapodes non mammaliens, secondairement bipèdes (certains Dinosauriens, Oiseaux).
- Ainsi, la tête de l’ulna, en position interne au niveau du coude, se retrouve en position externe au niveau du poignet, et inversement pour le radius.
- Membres : Les Grands Articles d'Universalis, Encyclopaedia Universalis, (lire en ligne), n.p..
- Les membres ancestraux des tétrapodes terrestres sont de type transversal (membres qui se plient en Z chez les Amphibiens du Primaire, les Urodèles et Reptiles actuels).
- Parallèlement, l'allongement des tendons permet de stocker plus d'énergie utile à la course quadrupède.
- André Beaumont, Pierre Cassier, Daniel Richard, Biologie animale. Les Cordés, Dunod, , p. 210
- Neil Shubin, Infographie montrant le passage de la nageoire des sarcoptérygyens tétrapodomorphes au membre chiridien des hommes, 3 février 2016.
- (en) Researchers say ability to throw played a key role in human evolution, phys.org (article initial dans Nature).
- Aurélien Miralles, « Sans pattes : quand les Tétrapodes ne le sont plus vraiment… », Espèces, no 19, , p. 24.
- En rouge le basipode, en violet le métapode, en jaune l'acropode, en marron les phanères terminales kératinisées. Le tarse correspond à la région du jarret, la pointe du jarret est homologue du talon.
{kind=link}
Voir aussi
[modifier | modifier le code]Bibliographie
[modifier | modifier le code]- (en) Sébastien Steyer, « Limbs:How do they work ? », dans Earth Before the Dinosaurs, Indiana University Press, (lire en ligne), p. 43-58
- (en) Neil Shubin, « Getting a grip », dans Your Inner Fish. A Journey Into the 3.5-Billion-Year History of the Human Body, Vintage Books,, (lire en ligne), p. 28-43
- (en) Brian K. Hall, Fins Into Limbs. Evolution, Development, and Transformation, University of Chicago Press, , 344 p. (lire en ligne)
Articles connexes
[modifier | modifier le code]- Chordés : anatomie comparée des vertébrés (André Baumont et Pierre Cassier).
- Pentadactylie
- Pronation et supination
- Membre ptérygien, autre type de membre chez les vertébrés