
Conceptacle algal



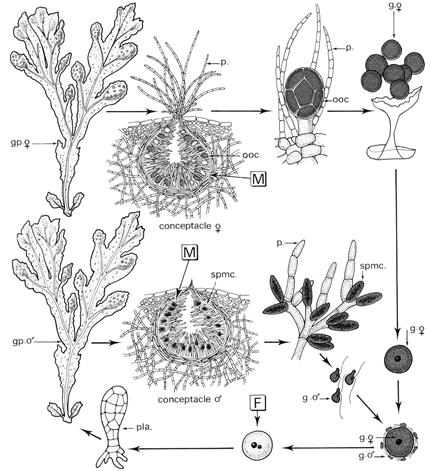
Un conceptacle algal est une cavité spécialisée construite dans le thalle de certaines algues marines, de milieux saumâtres ou d'eau douce, et qui abrite les cellules reproductrices. Les conceptacles algaux sont des cryptes pilifères fertiles (regroupées dans une portion du thalle appelée réceptacle) qui s'ouvrent le milieu extérieur par un petit orifice dénommé ostiole[5]. Des conceptacles sont présents chez les Corallinaceae[6], et Hildenbrandiales, ainsi que chez les algues brunes de l’ordre des Fucales. Chez les Fucales il n'y a pas de phase haploïde dans le cycle de reproduction et, par conséquent, aucune alternance des générations[7]. Les plantes diploïdes produisent des organes mâles (anthéridies) et femelles (oogonia) par méiose. Les gamètes sont libérés dans la mer. La fusion de deux gamètes planctoniques forme un zygote dont la division asymétrique génère une petite cellule qui se développe en un rhizoïde se fixant au substrat, et une grande cellule produisant le thalle[8].
Deux taxons, les corallines et les Hildenbrandiaceae, présentent des conceptacles mais qui ne sont pas homologues[9].
Des structures assez similaires existent aussi : les cryptes pilifères stériles, cavités réparties sur l'ensemble du thalle et qui ressemblent à des conceptacles, mais qui en diffèrent en ce qu'ils n'abritent que des poils et ne sont pas fertiles. Ces cryptes contribuent à la résistance des algues à la dessiccation lors de l'exondation en limitant les pertes d'eau liées à la transpiration[8].
Morphologie[modifier | modifier le code]
Les conceptacles peuvent mesurer 40 à 150 µm de diamètre chez certains Hildenbrandiales[10]. Certains conceptacles sont des trous bien visibles dans le thalle, tandis que d'autres sont protégés dans un dôme qui apparaît en saillie à la surface de l'algue[11]. Certains conceptacles s'ouvrent sur l'environnement aquatique de l'algue par le biais de plusieurs pores, les ostioles, qui diffèrent selon l'espèce et le type de conceptacle[11].
Chez l'algue brune Fucus distichus, les ostioles sont bloqués au cours de leur développement[5].
Composants[modifier | modifier le code]
- Columelle.
C'est le pilier central constitué de cellules calcifiées recouverts de débris cellulaires[11]. Ces cellules stériles se dressent sur le conceptacle sol pour former une pointe[12].
- Filaments ("roof filaments" pour les anglophones).
Certains conceptacles croissent de manière centripète en formant un trou près de la surface du thalle ; dans de tels cas, une sorte de toit constitué de filaments peut se former au-dessus du trou (filaments souvent long de 1 à 9 cellules)[13],[14] couvrant la chambre mais en laissant un pore central à travers lequel les spores pourront s'échapper[15]. Ces filaments peuvent aussi produire une ouverture en forme de bec[16].
- Spores.
Dans les conceptacles asexués les spores tendent à être de grande taille. Ils sont alors coincés dans la chambre du conceptacle dont il emplissent tout l'espace "comme les quartiers d'une d'orange"[11].
Types de conceptacle[modifier | modifier le code]
Il existe différents types de conceptacles, classés selon la nature des spores qu'ils contiennent ; certaines espèces peuvent en posséder jusqu'à quatre types distincts. Par exemple, Lithophyllum incrustans porte des conceptacles asexués, mâles, femelles et cystocarpiques (strictement, un type femelle) conceptacles, discuté ci-dessous[11].
- Conceptacles asexués.
Ils se développent là où des cellules faiblement calcifiées s'effondrent pour produire une cavité. Chez L. incrustans, leur columelle est centrale et bien visible (voir ci-dessus) ; il est accentué par les spores (bispores, tétraspores, etc.), qui se collent à la columelle et aux parois du conceptacle[11]. Un conceptacle asexué peut être unipore ou multipore[11].
- Conceptacles sexués (mâle/ femelle).
Ils n'ont pas de columelle[11]. L. incrustans présente des individus mâles et femelles distinct ; les deux types de conceptacle ne coexistent jamais sur le même thalle[11]. Les conceptacles mâles ont tendance à ne pas s'ancrer aussi profondément dans le thalle que les conceptacles femelles ; Leur taille est similaire, bien que leur forme diffère légèrement[11]. Les Cellules de la zone circulaire au sein du périthalle s'allongent pour former les conceptacles sexuels[11], laissant une trame de longues cellules autour de l'enceinte du conceptacle[17]. Dans le conceptacles mâle de l'algue Austrolithon, des filaments grandissent plus rapidement sur la marge, s'infléchissent sur le haut de la cavité du conceptacle pour éventuellement former un toit[15].
- Conceptacles cystocarpiques.
Ils ne sont pas associés à une déminéralisation de tissus mais commencent plutôt à se former à partir du centre (comme un conceptacle femelle) et ils se développent de manière radiale[11]. Comme les conceptacles femelles mûrs, il leur manque une columelle[11].
Développement[modifier | modifier le code]
Chez la plupart des algues du groupe des corallines, un amas de cellules reproductrices apparait dans la couche moyenne de l'algue, formant le conceptacle, lequel est englouti par les tissus environnants qui croissent pour produire un toit muni d'un unique pore au-dessus du conceptacle. Un bouchon peut même être ensuite produit pour en protéger l'ouverture[16]. Toutefois, le conceptacle peut généralement se former à n'importe quelle profondeur dans le thalle (dans la couche de surface ou en profondeur à la base du périthalle[11].
Quatre différents modes de formation de conceptacle asexué sont connus chez Corallina et Bossiella.
Chez les algues Bossiella (du groupe des corallines), le conceptacle se forme dans le cortex (la couche externe). Un épaississement se forme, avec séparation de l'epithallium externe de la couche sous-jacente du cortex ; la partie qui se soulève formera le chapeau du conceptacle alors que les cellules reproductrices grandissent en dessous[16]. une fois le bouchon formé, les cellules filamenteuses sous-jacentes se dégradent et les cellules du tissu qui entourent le chapeau se développent plus rapidement. Cette combinaison produit une chambre[16].
Histoire évolutive[modifier | modifier le code]
Des conceptacles apparaissent dans les archives fossiles, au moins dès le Silurien.
Des structures similaires ont été observées dans les Prototaxites, ce qui impliquerait que ce géant des terres émergées n'était pas un simple champignon (comme la plupart des paléontologues le supposent encore aujourd'hui), mais un grand lichen[18].
Notes et références[modifier | modifier le code]
- Nathalie Bourgougnon, Annette Gervois, Les algues marines. Biologie, écologie et utilisation, Ellipses, , p. 120.
- Cas fréquent chez les Fucales : chaque oogone, portée par un pédicelle, est formée initialement par une cellule unique qui se divise, ce qui génère huit oosphères (gamète femelles polyédriques) chargées de réserves. À marée basse, sous l'effet de la dessiccation, les gamètes perlent à l'extérieur des conceptacles. L'immersion provoque la rupture des enveloppes protectrices des gamètes et leur libération à travers les ostioles. Les oosphères sédimentent en milieu calme et les anthérozoïdes se dirigent vers le fond grâce au stigma (phototactisme négatif) et aux phéromones libérées par les oosphères (chimiotactisme positif à l'égard du fucoserratène chimiotactique des fucus).
- On observe aussi des poils stériles appelés paraphyses, non ramifiés.
- La richesse des algues en polysaccharides pariétaux (polysaccharides sulfatés formant une matrice anionique qui offre une surface d’hydratation importante et une certaine souplesse) leur permet de faire face à la dessiccation ou au stress mécanique (vagues, houle). Cf (en) Elizabeth Ficko-Blean, Cécile Hervé, Gurvan Michel, « Sweet and sour sugars from the sea: the biosynthesis and remodeling of sulfated cell wall polysaccharides from marine macroalgae », Perspectives in Phycology, vol. 2, no 1, , p. 51-64 (DOI 10.1127/pip/2015/0028).
- Boney, A.D. (1969).
- Irvine, L.M. and Chamberlain, Y.M. (1994).
- Fritsch, F.E. (1945).
- Flecher, R.L. (1987).
- Pueschel et Cole, « Rhodophycean Pit Plugs: an Ultrastructural Survey with Taxonomic Implications », American Journal of Botany, vol. 69, no 5, , p. 703–720 (DOI 10.2307/2442960, JSTOR 2442960)
- A. R. Sherwood et R. G. Sheath, « Systematics of the Hildenbrandiales (Rhodophyta): Gene Sequence and Morphometric Analyses of Global Collections », Journal of Phycology, vol. 39, , p. 409–422 (DOI 10.1046/j.1529-8817.2003.01050.x)
- R. G. J. Edyvean et B. L. Moss, « Conceptacle Development in Lithophyllum incrustans Philippi (Rhodophyta, Corallinaceae) », Botanica Marina, vol. 27, no 9, , p. 391–400 (DOI 10.1515/botm.1984.27.9.391)
- H. W. Johansen, « REPRODUCTION OF THE ARTICULATED CORALLINE AMPHIROA EPHEDRAEA », Journal of Phycology, vol. 4, no 4, , p. 319–328 (DOI 10.1111/j.1529-8817.1968.tb04702.x)
- K. Wilks et W. Woelkerling, « An account of southern Australian species of Phymatolithon (Corallinaceae, Rhodophyta) with comments on Leptophytum », Australian Systematic Botany, vol. 7, no 3, , p. 183 (DOI 10.1071/SB9940183)
- Roberta A. Townsend et John M. Huisman, « Epulo multipedes gen. et sp. nov. (Corallinaceae, Rhodophyta), a coralline parasite from Australia », Phycologia, vol. 43, no 3, , p. 288–295 (DOI 10.2216/i0031-8884-43-3-288.1, lire en ligne)
- A. S. Harvey et W. J. Woelkerling, « An account of Austrolithon intumescens gen. Et sp. Nov. And Boreolithon van-heurckii (Heydrich) gen. Et comb. Nov. (Austrolithoideae subfam. Nov., Corallinaceae, Rhodophyta) », Phycologia, vol. 34, no 5, , p. 362–382 (DOI 10.2216/i0031-8884-34-5-362.1)
- Johansen, H. W., « Ontogeny of Sexual Conceptacles in a Species of Bossiella (Corallinaceae) », Journal of Phycology, vol. 9, no 2, , p. 141 (DOI 10.1111/j.0022-3646.1973.00141.x)
- W. Woelkerling et A. Harvey, « An account of Southern Australian Species of Mesophyllum (Corallinaceae, Rhodophyta) », Australian Systematic Botany, vol. 6, no 6, , p. 571 (DOI 10.1071/SB9930571)
- J. M. Schopf, « Foerstia and recent interpretations of early, vascular land plants », Lethaia, vol. 11, no 2, , p. 139–143 (DOI 10.1111/j.1502-3931.1978.tb01298.x)
{kind=link}