
Architecture végétale

L'architecture végétale est une discipline botanique fondée sur l'analyse morphologique qui cherche à rendre compte de l'organisation spatio-temporelle de l'appareil végétatif des plantes. Cette architecture est, à tout moment, l'expression d'un équilibre entre des processus endogènes de croissance (croissance primaire en longueur et secondaire en diamètre, ramification intrinsèque et réitération, processus sous fort contrôle génétique) et des contraintes extérieures exercées par le milieu (sol, climat, compétition intra et interspécifique, taille…)[4].
Elle désigne par extension l'organisation fondamentale d'un végétal, déterminée génétiquement et construite par le libre fonctionnement des méristèmes aériens (rameaux reproducteurs et caulinaires, ces derniers imprimant l'aspect général à la plante) et souterrains (ramification racinaire)[5],[6].
Historique[modifier | modifier le code]
Le terme « architecture végétale » est proposé par le botaniste britannique Edred John Henry Corner (1906-1995) dans les années 1940 pour définir la morphologie des structures aériennes des arbres.
Les travaux de Francis Hallé[7] et Oldeman décrivent en 1970 la mise en place des axes végétatifs pendant le développement des arbres des forêts tropicales. Ils effectuent une mise à jour des données sur l'architecture végétale et établissent une typologie selon des modèles architecturaux arborescents basés sur les modes de ramification des arbres, la rythmicité de ces ramifications, leur dynamique et leur position[8]. Les recherches à leur suite qui étudient le développement des axes végétatifs des plantes permet d'identifier une vingtaine de modèles communs aux végétaux vasculaires terrestres.
L'approche de Hallé et Oldeman n'échappe aux écueils d'une modélisation réductrice[9]. Les modèles architecturaux des plantes qui appartiennent à un groupe évolutionnaire tardif, sont en effet sous la dépendance de nombreux autres facteurs (par exemple l'inhibition du développement d'une branche par une autre branche, due à la fois à des signaux internes et à l'ombrage mutuel)[10].
L'analyse architecturale est un domaine de recherche actif qui devient pluri-disciplinaire à partir des années 2000, grâce aux progrès de l'informatique qui permettent de concevoir des modèles « structure-fonction » (FSPM) ou « plantes virtuelles » adaptés au sujet : arbres combinatoires (Janey 1992[11]), grammaires (Pruzinkiewicz, 1988[12], Kurth 1989[13]), fractals (Smith, 1984)[14], système multi-agents (Eschenbach, 2005)[15], etc[16].
Concepts en architecture végétale[modifier | modifier le code]


Au niveau de l'anatomie des architectures végétales, toutes les plantes vasculaires ont en commun les mêmes composants de base architecturaux : le système aérien, appareil caulinaire composé d'axes végétatifs (ou axes feuillés car ce sont le plus des tiges feuillées) ramifiés et le système racinaire, séparés par un collet. L’axe végétatif est édifié à partir d'un méristème apical caulinaire et racinaire qui mettent en place par allongement des séries de phytomères, unités botaniques fonctionnelles qui s'empilent et portent les organes, éléments de base de l’architecture végétale. La disposition de ces organes le long des axes s’ordonne selon les règles de la phyllotaxie. Les axes végétatifs se ramifient pour constituer une structure végétale dans laquelle on peut identifier des ordres de ramifications (l’ordre 1 est la tige principale, l’ordre 2 les rameaux secondaires, l'ordre 3 les rameaux de troisième ordre, etc.). Plusieurs axes végétatifs disposés selon un ordre hiérarchisé sont désignés par le terme d'unité architecturale[18]. Ce concept d'axe végétatif et de catégorie d'axes, qui s'applique aussi bien au niveau des tiges (catégories caulinaires)[19] que des racines (catégories racinaires)[20], s'est d'abord développé dans le contexte d'études menées sur des arbres des zones tropicales, avant qu'elles ne soient adaptées à ceux des zones tempérées[21].
La réitération est le processus par lequel un organisme duplique totalement ou partiellement sa propre architecture (clonage par reproduction asexuée, à l'origine de nouveaux individus appelés ramets). Ce duplicate est appelé réitérat. La réitération séquentielle met en place des syllepses (appelés aussi rameaux sylleptiques, réitérats immédiats ou séquentiels). La réitération différée ou retardée donne des prolepses (appelés aussi rameaux proleptiques, réitérats différés ou retardés). « Cette prolifération de termes, souvent gênante, est le reflet de la lente évolution des idées[22] ». Les unités architecturales sont des réitérats qui deviennent, d'année en année, de plus en plus petits et de plus en plus nombreux en se développant à la périphérie du houppier. Chaque unité correspond, selon le degré de complexité, à la différenciation d'un nombre défini de catégories d'axes différentes (cinq chez le platane). Au fur et à mesure que la ramification progresse à la périphérie de l'arbre, il se développe ainsi des séries de rameaux généralement plus simples (les rameaux florifères).
Méthode[modifier | modifier le code]
Le modèle architectural d'une plante est défini par :
- le mode de ramification des axes : absent ou présent (ramification sympodiale ou monopodiale) ;
- le mode de croissance des axes : continue ou rythmique ;
- l'orientation des axes secondaires : orthotropes ou plagiotropes ;
- la position de la sexualité : latérale ou terminale.
Intérêts des modèles architecturaux[modifier | modifier le code]
« La connaissance des traits morphologiques qui concernent la structure globale de la plante et qui sont impliqués dans le passage d’une forme de croissance à une autre est fondamentale en biologie ; notamment pour des aspects taxonomiques, évolutifs, agronomiques et écologiques. Si ces traits sont plastiques, ils peuvent permettre d’estimer la fitness des individus d’une population, notamment au travers de la caractérisation des stratégies d’occupation de l’espace et des fonctions développementales (Bradshaw, 1965[23] ; Briggs et Walters, 1997[24] ; Zhukova, 2001[25] ; Zhukova et Glotov, 2001[26] ; Pérez-Harguindeguy et al., 2013[27]). Si ces traits ne sont pas plastiques mais variables en relation à l’appartenance à un taxon de rang inférieur (sous-espèce, variété, écotype, chémotypes, etc.), ils reflètent alors des mécanismes potentiellement adaptatifs et révèlent les relations morphologiques qui existent entre les différentes formes de croissance, ouvrant la voie à la compréhension de leur phylogenèse (Troll, 1937[28] ; Rauh, 1962[29])[30] ».
Les études en architecture végétale appliquées à la biologie peuvent servir à proposer des voies possibles dans l'évolution des formes dans un même phylum. En agronomie, elles sont utilisées pour établir des modélisations du rendement des plantes cultivées influencé par de nombreux facteurs sur la structure de l'arbre (attaque de bioagresseurs, produits phytosanitaires, engrais, amendements, verse, densité de la population végétale)[31].
Modèles architecturaux[modifier | modifier le code]
Chaque modèle architectural a été nommé en référence à un botaniste.
Hallé et Oldeman ont recensé 24 modèles dans le monde végétal[32].
- Holttum : un axe unique (monocaule) orthotrope, avec des organes sexuels terminaux qui provoquent la mort de la plante. C'est le cas du bananier, des agaves et des palmiers monocarpiques comme le talipot. Ce modèle a été nommé en référence au botaniste Richard Eric Holttum pour ses travaux sur les caractéristiques limitantes de la croissance des monocotylédones où il décrit des formes qui ne fleurissent qu'une seule fois avant de mourir.
-

-
 Agave sp.
Agave sp. -
 Grande bardane (Arctium lappa)
Grande bardane (Arctium lappa) -

-
 Tournesol
Tournesol





- Corner : un seul axe (monocaule) orthotrope à sexualité latérale qui permet une croissance indéfinie de l'axe végétatif. Le Cycas revoluta femelle, de nombreux palmiers, le palmier à huile et le papayer font partie des nombreux représentants de ce modèle. Les arbres qui suivent ce modèle n'ont pas de branches, mais l'axe unique peut chez certaines espèces porter des réitérats : ainsi, les apparentes « branches » du papayer sont en réalité des reproductions en plus petit de l'axe principal.





- Tomlinson : répétition de modules orthotropes à partir d'une ramification basale (polycaule) à sexualité terminale ou latérale. C'est ce modèle que suivent les palmiers cespiteux.
- Chamberlain : tronc constitué d'une succession subapicale de modules orthotropes verticaux à floraison terminale. La plante apparaît comme monocaule mais est en fait un sympode linéaire monochasial. L'arbre corail (Jatropha multifida) et le Cycas revoluta mâle est expriment ce modèle.
-
 Floraison terminale de Jatropha multifida.
Floraison terminale de Jatropha multifida. -
 Inflorescence du Cycas revoluta mâle.
Inflorescence du Cycas revoluta mâle.


- Leeuwenberg : sympode à floraison terminale formé d'une succession de modules orthotropes à ramification subapicale. Contrairement au modèle de Koriba, les axes résultant de la fourchaison restent tous orthotropes (pas de changement de fonction d'un des axes devenant plagiotrope pour assurer le rôle d'une branche). Ce modèle est fréquent chez les Apocynacées (ramification binaire chez le frangipanier blanc, ternaire chez le laurier-rose), les Solanacées, les Euphorbiacées (Ricin commun ricin, sumacs)[33].
-
 Ramification dichotomique du frangipanier blanc.
Ramification dichotomique du frangipanier blanc.

- Schoute : succession de modules dichotomiques à ramification terminale (sans mort d'apex) et sexualité latérale. C'est un modèle peu répandu, représenté notamment par le remarquable palmier doum.
- Koriba : succession de sympodes modulaires redressés, relais subterminaux, branches plagiotropes par apposition, sexualité terminale. Ailanthus altissima suit ce modèle de développement[34].
- Prévost : tronc sympodial, modules orthotropes à développement différé, branches sympodiales plagiotropes par apposition de modules à développement immédiat, sexualité terminale.
- Fagerlind : tronc orthotrope à ramification rythmique. Les axes secondaires sont des modules sympodiaux à sexualité terminale portée par l'extrémité orthotrope et dont la base est plagiotrope par apposition. L'implantation des feuilles (phyllotaxie) est spiralée. On retrouve ce modèle chez les magnolias.
- Aubréville : tronc monopodial orthotrope à ramification rythmique. Les axes secondaires sont des modules sympodiaux plagiotropes par apposition (base plagiotrope et partie distale orthotrope), sexualité latérale. Ce modèle est celui du badamier.
-
 Modules sympodiaux plagiotropes par apposition formant les branches de Terminalia catappa
Modules sympodiaux plagiotropes par apposition formant les branches de Terminalia catappa -
 Fructification latérale de Terminalia catappa
Fructification latérale de Terminalia catappa


- Petit : tronc monopodial orthotrope, ramification continue, branches sympodiales, sexualité terminale.
- Scarrone : tronc monopodial orthotrope à ramification rythmique. Les branches orthotropes sont des modules sympodiaux à sexualité terminale. C'est le modèle de développement du manguier et du marronnier d'Inde.
-
 Aesculus hippocastanum en hiver
Aesculus hippocastanum en hiver -
 Floraison terminale d'Aesculus hippocastanum
Floraison terminale d'Aesculus hippocastanum -

-
 Floraison terminale de Mangifera indica
Floraison terminale de Mangifera indica


;_fruiting_tree,_leaves,_fruit,_f_Wellcome_V0042597.jpg)

- Stone : monopode orthotrope à ramification continue ou diffuse, branches orthotropes sympodiales à sexualité terminale.
- Rauh : monopode à croissance rythmique, tronc et branches orthotropes et sexualité latérale. Les espèces pionnières des zones tropicales (hévéa, Cecropia, certaines espèces de chênes et d'érables) ou tempérées (pins, peupliers, frênes sauf Fraxinus ornus qui a une floraison terminale), semi-pionnières (noyer commun)[35] présentent une architecture conforme à ce modèle[33],[36].
- Attims : monopode à croissance continue, tronc et branches orthotropes et sexualité latérale. Le palétuvier blanc, Alnus glutinosa et le cyprès de Provence obéissent à ce modèle. C'est un modèle proche de celui de Rauh, sauf la croissance qui est continue au lieu de rythmique (ce qui induit une ramification diffuse et non rythmique).
- Nozeran : tronc sympodial orthotrope composé de modules à développement différé, branches monopodiales plagiotropes, sexualité latérale.
- Massart : monopode à croissance rythmique, tronc orthotrope, branches plagiotropes, sexualité latérale ou terminale. C'est le modèle de développement du platane[37], des frênes, de Taxus baccata, de Sequoia sempervirens, de Pseudotsuga menziesii et du Ginkgo biloba.
-
 Taxus baccata laissant apparaitre un tronc orthotrope autour duquel s'organisent des axes secondaires plagiotropes.
Taxus baccata laissant apparaitre un tronc orthotrope autour duquel s'organisent des axes secondaires plagiotropes. -
 Axes secondaires sur le tronc d'un Ginkgo biloba traduisant une croissance rythmique.
Axes secondaires sur le tronc d'un Ginkgo biloba traduisant une croissance rythmique. -
 Axes principaux orthotropes et axes secondaires plagitropes d'un Ginkgo biloba.
Axes principaux orthotropes et axes secondaires plagitropes d'un Ginkgo biloba. -
 Axe principal orthotrope (tronc) et axes secondaires plagitropes (branches) de Pseudotsuga menziesii
Axe principal orthotrope (tronc) et axes secondaires plagitropes (branches) de Pseudotsuga menziesii -
 Le tronc vertical orthotrope de Sequoia sempervirens peut dépasser 100 m de haut. En revanche, ses branches plagiotropes plongent en direction du sol.
Le tronc vertical orthotrope de Sequoia sempervirens peut dépasser 100 m de haut. En revanche, ses branches plagiotropes plongent en direction du sol. -
 Exempt de tailles de mise en forme et d'élagage, le platane commun arbore naturellement des branches plagiotropes.
Exempt de tailles de mise en forme et d'élagage, le platane commun arbore naturellement des branches plagiotropes.



.jpg)


- Roux : monopode à croissance continue ou diffuse, tronc orthotrope, branches plagiotropes, sexualité essentiellement latérale. C'est le modèle de développement du caféier d'Arabie. C'est un modèle proche de celui de Massart, sauf la croissance qui est continue au lieu de rythmique (ce qui induit une ramification diffuse et non rythmique).
-
 Axes secondaires de Coffea arabica se développant de manière rythmique de part et d'autre de l'axe principal.
Axes secondaires de Coffea arabica se développant de manière rythmique de part et d'autre de l'axe principal. -
 Branche plagiotrope et fructifications latérales de Coffea arabica traduisant une croissance rythmique.
Branche plagiotrope et fructifications latérales de Coffea arabica traduisant une croissance rythmique.


- Champagnat : sympode formé d'axe orthotropes s'affaissant sur la partie distale, les relais de croissance apparaissant dans la courbure. Rosa canina, Bougainvillea spectabilis et Rubus idaeus sont conformes à ce modèle.
-
 Axe orthotrope de Bougainvillea spectabilis avec apparition des relais de croissance dans la courbure.
Axe orthotrope de Bougainvillea spectabilis avec apparition des relais de croissance dans la courbure.
.JPG)
- Mangenot : superposition d'axes mixtes (orthotropes sur la partie proximale, plagiotropes sur la partie distale). C'est le modèle de développement de la myrtille américaine.
-
 Vaccinium corymbosum se dresse sur une succession d'axe orthotropes alors que la partie distale prend une forme plagiotrope.
Vaccinium corymbosum se dresse sur une succession d'axe orthotropes alors que la partie distale prend une forme plagiotrope.

- Troll : plagiotrope sur tous les axes, la constitution du tronc se fait par superposition et/ou redressement de la partie proximale d'un axe. Fagus grandifolia, Cercis siliquastrum,Corylus avellana[38] et Ulmus sont conformes à ce modèle.
-
 Cercis siliquastrum est constitué d'une superposition d'axes plagiotropes.
Cercis siliquastrum est constitué d'une superposition d'axes plagiotropes. -
 Ulmus americana ne laissant apparaitre qu'une structure plagiotrope.
Ulmus americana ne laissant apparaitre qu'une structure plagiotrope.


- Bell : production au départ d'un axe horizontal souterrain dont les ramifications latérales produisent des axes secondaires orthotropes aériens. Les axes secondaires aériens peuvent prendre l'aspect de n'importe quel autre modèle. Les bambous, en particulier à rhizome traçant, sont des représentants typiques de ce modèle.
Distribution et expression des modèles[modifier | modifier le code]
Les modèles de Rauh, Massart et Troll sont les plus courants aux hautes latitudes. Les zones équatoriales et inter-tropicales présentent une diversité de modèles beaucoup plus importante. Les modèles les plus courants présentent une plus grande diversité d'expression des caractères que les modèles plus rares. L'expression du modèle est souvent plus visible sur une jeune plantule que sur un arbre âgé.
Classement des modèles[modifier | modifier le code]
Les modèles peuvent être regroupés en six classes :
- Classe I (mono-axial) : Holttum, Corner ;
- Classe II (poly-axial) : Chamberlain, Tomlinson, Leeuwenberg, Schoute ;
- Classe III (sympode à axes différenciés) : Koriba, Nozeran, Prévost ;
- Classe IV (axes autodifférenciés) : Champagnat, Mangenot, Troll ;
- Classe V (monopode à axes différenciés) : Stone, Attims, Petit, Roux, Scaronne, Faguerlind, Rauh, Aubréville, Massart ;
- Classe VI (axe principal horizontal et axes secondaires orthotropes) : Bell.
Architecture végétale des conifères[modifier | modifier le code]
Les conifères connaissent quatre modèles de croissance[39] :
- Rauh : Pinus, Picea, Sequoia, Metasequoia, Cunnighamia, Cryptomeria, Callitris, Dacrydium, Neocalliptopsis, Agathis, Araucaria, Decussocarpus, Dacrycarpus, Austrotaxus.
- Massart : Abies, Agathis, Araucaria, Taxus, Cephallotaxus.
- Attims : Cupressus, Chamaecyparis, Thuya, Juniperus.
- Mangenot : Tsuga, Libocedrus, Acmopyle, Falcatifolium.
L'étude de l'architecture des conifères laisse apparaître des modulations architecturales.
Certains genres juxtaposent deux modèles sur le même arbre : à la phyllotaxie spiralée de la partie proximale des branches orthotropes de Sequoia sempervirens succède sur la partie distale des ramifications plagiotropes portant des limbes foliaires disposés dans le plan. Ainsi la partie basale de l'arbre est conforme au modèle de Rauh, la partie apicale à celui de Massart.
D'autres semblent développer des stratégies de croissance intermédiaires entre deux modèles :
- Picea, Rauh (tronc et axes principaux orthotropes) et Massart (axes secondaires plagiotropes) ;
- Chamaecyparis et Thuya, Attims (tronc et axes principaux monopodiaux orthotropes à croissance continue) et Massart (axes secondaires plagiotropes).
Enfin, d'autres changent de modèle au cours de leur croissance : c'est le cas des Araucarias et d’Abies passent de Massart à Rauh.
Étant donné le caractère archaïque des conifères, comparé aux angiospermes, leurs modulations architecturales pourraient traduire une restriction progressive dans l'expression des formes végétales qui aurait abouti aux modèles actuels.
Emboîtement des modèles[modifier | modifier le code]
En 2012, Jeanne Millet propose qu'à côté de l'expression strictement « conforme » à un modèle défini, plusieurs modèles peuvent apparaitre au sein d'un même arbre, témoignant de différents niveaux d'organisation et de hiérarchie[40].
La plantule initiale peut apparaitre comme un module « apparenté » à un modèle dit « simple », puis en se ramifiant va s'exprimer dans un modèle plus « complexe » qui conditionne la ramification et organise les réitérations.
En identifiant plusieurs unités architecturales au sein d'un même arbre, on peut alors parler modèles emboîtés :
- Leeuwenberg de Massart pour le platane commun, le hêtre commun, le châtaignier commun,
-
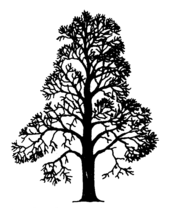 Le développement des réitérations du châtaignier semblent apparentées au modèle de Leeuwenberg
Le développement des réitérations du châtaignier semblent apparentées au modèle de Leeuwenberg -
 Les inflorescences et les axes secondaires du châtaignier obéissent au modèle de Massart
Les inflorescences et les axes secondaires du châtaignier obéissent au modèle de Massart -
 Les réitérations du platane apparentées au modèle de Leeuwenberg
Les réitérations du platane apparentées au modèle de Leeuwenberg -
 Les inflorescences et axes secondaires du platane conforment au modèle de Massart
Les inflorescences et axes secondaires du platane conforment au modèle de Massart -
 Les réitérations du hêtre commun (Fagus sylvatica) suivent le modèle de Leeuwenberg
Les réitérations du hêtre commun (Fagus sylvatica) suivent le modèle de Leeuwenberg -
 Modèle de Massart chez le hêtre
Modèle de Massart chez le hêtre
.png)





- Leeuwenberg de Rauh pour le noyer noir, le peuplier noir,
-
 Modèle de Leeuwenberg s'exprimant dans les réitèrations du noyer
Modèle de Leeuwenberg s'exprimant dans les réitèrations du noyer -
 Inflorescence du peuplier noir (Rauh)
Inflorescence du peuplier noir (Rauh)


- Leeuwenberg de Koriba de Rauh pour le chêne pédonculé : il passe successivement par les modèles de Holttum (unité minimale), Rauh, Koriba et Leeuwenberg, tous ces modèles étant présents à différentes échelles dans l'arbre adulte.
Notes et références[modifier | modifier le code]
- Appelé localement « Ar Ven Der » (« l'arbre chêne »), ce chêne pédonculé a un âge estimé entre 800 (selon Drénou) et 1200 ans (selon Ted Green (en)).
- Certaines charpentières comptent plus de 80 cernes, ce qui indique que la taille en têtard a été abandonnée au début du XXe siècle.
- Christophe Drénou, Face aux arbres : apprendre à les observer pour les comprendre, Ulmer, , p. 27
- Claude Édelin, « L'architecture monopodiale : l'exemple de quelques arbres d'Asie tropicale », Thèse de doctorat d'État, Univ. Montpellier II, 1984, 258 p.
- Les connaissances et mesures de l'architecture racinaire (ou architecture des systèmes racinaires : traçant, en cœur — oblique, pivotant, à pivots multiples), encore fragmentaires dans les années 1990, se développent dans les années 2000. Cf Coutts M.P., 1983. Root architecture and tree stability. Plant Soil 71, 171-188 ; Danjon F., Reubens B., 2008. Assessing and analyzing 3D architecture of woody root systems, a review of methods and applications in tree and soil stability, resource acquisition and allocation. Plant and Soil 303, 1-34.
- « À la germination, la graine émet une radicule (le pivot primaire) qui s'allonge très vite et dont la longueur, en fin de première année, excède fréquemment celle de la tige. Par la suite, d'autres axes analogues au pivot initial (les pivots secondaires) apparaîtront sous les grosses racines horizontales. Cet ensemble de pivots verticaux (et obliques chez certaines essences) constitue le réseau plongeant. Il dépasse rarement une profondeur de 1,50 m.
Le réseau traçant réunit les racines ligneuses portées par les pivots ou le collet et ayant une direction de croissance horizontale. Ces racines charpentières peuvent être particulièrement longues , souvent bien plus que l'arbre n'est haut ». Cf Christophe Drénou, Les racines. Face cachée des arbres, Institut pour le développement forestier, , p. 69 - « Ein Leben im Zeichen des Urwalds » (consulté le )
- F. Hallé, R. Oldeman, Essai sur l'architecture et la dynamique de croissance des arbres tropicaux, 1970, 178 p.
- (en) Tsvi Sachs & Ariel Novoplansky, « Tree form: architectural models do not suffice », Israel Journal of Plant Sciences, vol. 43, no 3, , p. 203-212 (DOI 10.1080/07929978.1995.10676605).
- (en) Franck Varenne, From Models to Simulations, Routledge, , p. 81
- Nicolas Janey, 1992. Modélisation et synthèse d'images d'arbres et de bassins fluviaux associant méthodes combinatoires et plongement automatique d'arbres et cartes planaires., thèse en automatique et informatique de l'université de Franche-Compté, 321 p.
- (en) P. Prusinkiewicz, A. Lindenmayer, J. Hanan, « Developmental Models of Herbaceous Plants for Computer Imagery Purposes », SIGGRAPH '88, Computer Graphics, vol. 22, no 4, , p. 141–150 (DOI 10.1145/54852.378503)
- (en) Winfried Kurth, « Morphological models of plant growth: Possibilities and ecological relevance », Ecological Modelling, vol. 75–76, , p. 299-308 (DOI 10.1016/0304-3800(94)90027-2)
- (en) Alvy Ray Smith, « Plants, fractals and formal languages », Computer Graphics, vol. 18, no 3, , p. 1–10 (lire en ligne)
- (en) Christiane A. Eschenbach, « Emergent properties modelled with the functional structural tree growth model ALMIS : Computer experiments on resource gain and use », Ecological Modelling, vol. 186, no 4, , p. 470-488 (DOI 10.1016/j.ecolmodel.2005.02.013)
- Philippe de Reffye, Marc Jaeger, Daniel Barthélémy, François Houllier, Architecture des plantes et production végétale. Les apports de la modélisation mathématique, éditions Quæ, (lire en ligne), p. 14-18
- La phase de sénescence commence par la dislocation lente du houppier qui perd ses branches maîtresses et se poursuit par la mort progressive des branches ultimes. Cette mortalité de la périphérie de l'arbre (précédée par une réduction des modules réitérés, une nanification et la formation d'une unité architecturale minimale en coraux au niveau de ses branches les plus hautes) est associée à une régénération partielle de sa couronne (réitérations proleptiques tardives de plus en plus importantes et plus proches de la base des charpentières, se développant sur leur face supérieure : passage à l'épitonie) avant que cette dernière ne soit touchée par une mortalité centrioète. Cf Barthélémy D., Caraglio Y., Drénou C., Figureau C., « Architecture et sénescence des arbres », Forêt Entreprise, no 83, , p. 15-35
- Philippe de Reffye, « Modélisation de la croissance des plantes. Cas du modèle GreenLab », Bulletin mensuel de la Société linnéenne de Lyon, vol. 86, nos 5-6, , p. 142-143
- Claude Edelin, Images de l'architecture des Conifères, Thèse Spécialité, Univ. Montpellier., 1977, 254p.
- Francis Kahn, « Analyse structurale des systèmes racinaires des plantes ligneuses de la forêt tropicale dense humide », Candollea, vol. 32, no 2, , p. 321-358
- (en) Amram Eshel et Tom Beeckman, Plant Roots : The Hidden Half, CRC Press, , p. 29-9 à 29-12
- Christophe Drénou, La taille des arbres d'ornement : du pourquoi au comment, Forêt privée française, , p. 253
- (en) Anthony David Bradshaw, « Evolutionary Significance of Phenotypic Plasticity in Plants », Advances in Genetics, vol. 13, , p. 115-155 (DOI 10.1016/S0065-2660(08)60048-6).
- (en) David Briggs et Stuart Max Walters, Plant variation and evolution, Cambridge University Press, , 512 p. (lire en ligne)
- (en) L. A. Zhukova, « Diversity of Ontogenetic Pathways in Plant Populations », Russian Journal of Ecology, vol. 32, , p. 151–158 (DOI 10.1023/A:1011301909245)
- (en) L. A. Zhukova & N.V Glotov, « Morphological polyvariance of ontogeny in natural 692 plant populations », Russian Journal of Developmental Biology, vol. 32, , p. 381-387 (DOI 10.1023/A:1012838120823)
- (en) N. Pérez-Harguindeguy et al., « New handbook for standardised measurement of plant 641 functional traits worldwide », Australian Journal of Botany, vol. 61, , p. 167-234 (DOI 10.1071/BT12225)
- (de) Wilhelm Troll, Vergleichende morphologie der höheren pflanzen, Gebrüder Borntraeger, Koenigstein-Taunus, .
- (de) Werner Rauh, « Bemerkenswerte Sukkulente aus Madagaskar-12. Die Pachypodium-Arten 645 Madagaskars », Kakteen und andere Sukkulenten, vol. 13, no 3, , p. 96-103
- Mathieu Millan. Analyse de la variabilité des traits architecturaux des formes de croissance dans les communautés végétales. Botanique. Université de Montpellier, 2016, p. 40
- Claude Evelin, L'arbre, Naturalia Monspeliensia, , p. 224
- Franck Varenne, Du modèle à la simulation informatique, Vrin, , p. 203
- (en) Lauritz B. Holm-Nielsen, Ivan Nielsen, Henrik Balslev, Tropical forests : botanical dynamics, speciation and diversity, Academic Press, , p. 90
- (en) Ingo Kowarik, Ina Säumel, Biological flora of Central Europe: Ailanthus altissima (2007) Lire en ligne
- Yves Caraglio, Claude Édelin, Le Platane, quelques aspects de son architecture Lire en ligne.
- (en) F. Halle, R.A.A. Oldeman, P.B. Tomlinson, Tropical Trees and Forests : An Architectural Analysis, Springer Science & Business Media, , p. 221
- Yves Caraglio, Pascal Dabadie, Le Peuplier, quelques aspects de son architecture Lire en ligne.
- A. Schnitzler, L’intérêt du modèle architectural dans l’analyse de la biodiversité forestière. Application à la gestion des réserves naturelles rhénanes , Rev. For. Fr. LIII numéro spécial 2001.Lire en ligne
- Claude Edelin, 1977-1980
- Jeanne Millet, L'Architecture des arbres des régions tempérées, Éd. Multimonde, 2012, p. 77-81.
Annexes[modifier | modifier le code]
Bibliographie[modifier | modifier le code]
- C. Édelin, « Quelques aspects de l'architecture végétative des Conifères » in Bulletin de la Société Botanique de France, Lettres Botaniques, 128:3, 177-188 Lire en ligne.
- C. Édelin, Réflexions sur la compartimentation et l'intégration de l'architecture des arbres, 1997. Lire en ligne
- F. Hallé, R. A. A. Oldeman et P. B. Tomlinson, Tropical trees and forest An Architectural Analysis. Springer Verlag, Berlin. 1978, 441 p. Extraits en ligne
- F. Hallé, Plaidoyer pour l'arbre, Acte Sud, 2005.
- J. Millet, L'Architecture des arbres des régions tempérées, Ed. Multimonde, 2012. Extrait en ligne
Articles connexes[modifier | modifier le code]
Lien externe[modifier | modifier le code]
- Les 23 modèles théoriques en images sur fr.slideshare.net